
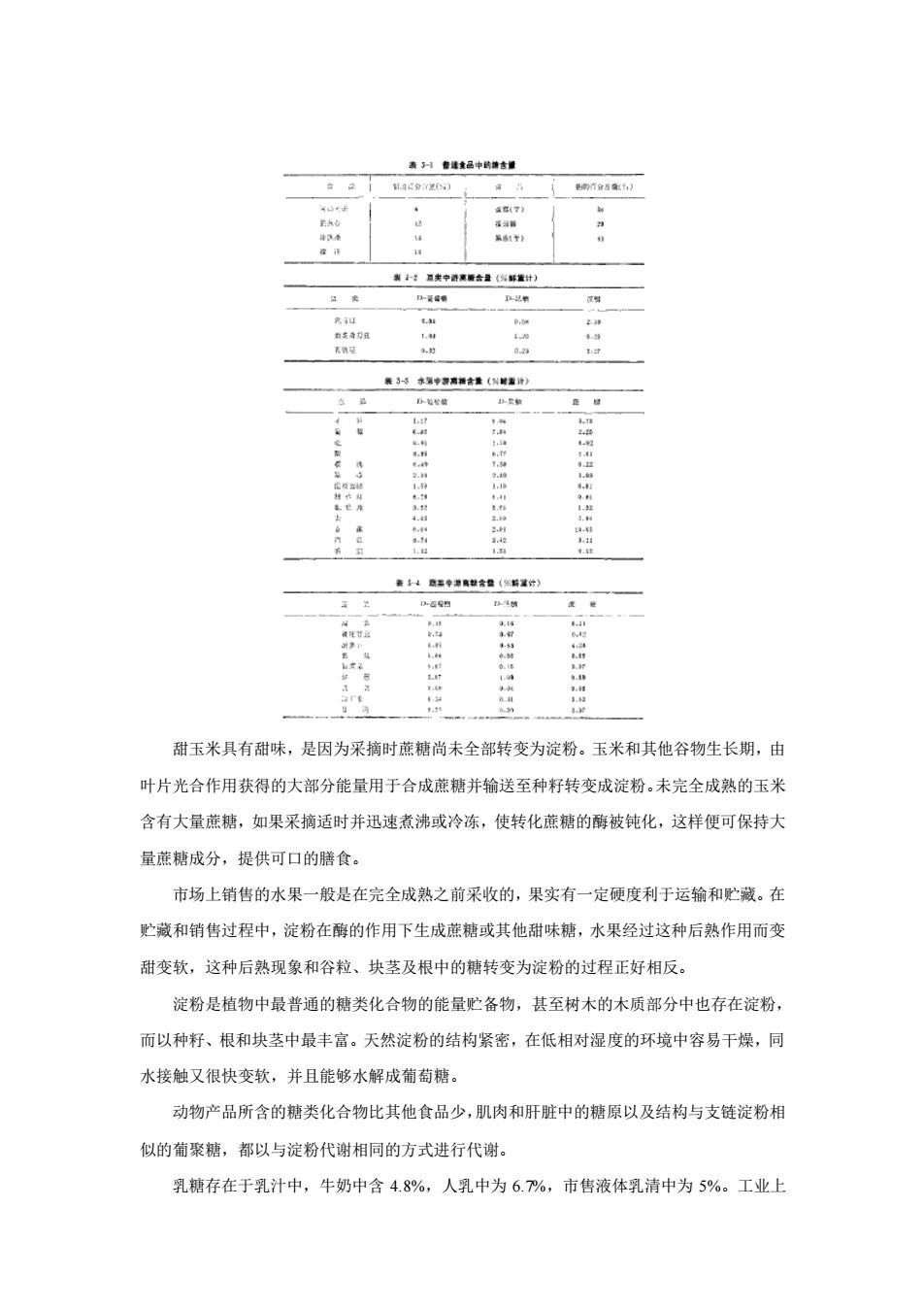
第三章 糖 类 第一节 概述 糖类化合物是自然界分布广泛、数量最多的有机化合物,是绿色植物光合作用的直接产 物。自然界的生物物质中,糖类化合物约占四分之三,从细菌到高等动物都含有糖类化合物, 植物休中含量最丰富,约占其干重的 85~90%,其中以纤维素最为丰富。其次是节肢动物, 如昆虫,蟹虾外壳的壳多糖(甲壳质)。 糖类化合物的分子组成可用 Cn(H2O) m的通式表示,统称为碳水化合物。但后来发现有 些糖如鼠李精(C6H12O5)和脱氧核糖 (C5H10O4)并不符合上述通式,并且有些糖还含有氮、硫、 磷等成分,显然碳水化合物的名称已经不适当,但由于沿用已久,至今还在使用这个名词。 根据糖类的化学结构特征,糖类的定义应是多羟基醛或酮及其衍生物和缩合物。 糖类化合物是生:物体维持生命活动所需能量的主要来源,是合成其他化合物的基本原 料,同时也是生物体的主要结构成分。人类摄取食物的总能量中大约 80%由糖类提供,因 此,它是人类及动物的生命源泉。我国传统膳食习惯是以富含糖类化合物的食物为主食,但 近年来随着动物蛋白质食物产量逐年增加和食品工业的发展,膳食的结构也在逐渐地变化。 一、糖类化合物的种类 糖类化合物的分类常按其组成分为单糖,寡糖和多糖。单糖是一类结构最简单的糖。是 不能再被水解的糖单位。根据其所含原子的数目分为丙糖、丁糖、戊糖和己糖等。根据官能 团的特点又分为醛糖或酮糖。寡糖一般是由 2~10 个分子单糖缩合而成,水解后产生单糖。 多糖是由多个单糖分子缩合而成,多糖中由相同的单糖基组成的称同聚多糖,不相同的单糖 基组成的称杂聚多糖,如按其分子中有无支链,则有直链、支链多糖之分。按其功能不同, 则可分为结构多糖、贮存多糖,抗原多糖等。 二、食品中的糖类化合物 陆地植物和海藻干重的四分之三由糖类化合物构成。谷物、疏菜、果实和可供食用的其 他植物都含有糖类化合物。 大多数植物只含少量蔗糖,大量膳食蔗糖来自经过加工的食品。在加工食品中添加的蔗 糖量一般是比较多的(表 3-1)。蔗糖是从甜菜或甘蔗中分离得到的,果实和蔬菜中只含少量 蔗糖、D-葡萄糖和 D-果糖(表 3-2,表 3-3 和表 3-4)。 谷物只含少量的游离糖,大部分游离糖榆送至种籽中并转变为淀粉,玉米粒含 0.2%~ 0.5%D-葡萄糖、0.1~0.4%D-果糖和 1~2%的蔗糖,小麦粒中这几种糖的含量分别小于 0.1%, 0.1%和 1%
第三章 糖 类 第一节 概述 糖类化合物是自然界分布广泛、数量最多的有机化合物,是绿色植物光合作用的直接产 物。自然界的生物物质中,糖类化合物约占四分之三,从细菌到高等动物都含有糖类化合物, 植物休中含量最丰富,约占其干重的 85~90%,其中以纤维素最为丰富。其次是节肢动物, 如昆虫,蟹虾外壳的壳多糖(甲壳质)。 糖类化合物的分子组成可用 Cn(H2O) m的通式表示,统称为碳水化合物。但后来发现有 些糖如鼠李精(C6H12O5)和脱氧核糖 (C5H10O4)并不符合上述通式,并且有些糖还含有氮、硫、 磷等成分,显然碳水化合物的名称已经不适当,但由于沿用已久,至今还在使用这个名词。 根据糖类的化学结构特征,糖类的定义应是多羟基醛或酮及其衍生物和缩合物。 糖类化合物是生:物体维持生命活动所需能量的主要来源,是合成其他化合物的基本原 料,同时也是生物体的主要结构成分。人类摄取食物的总能量中大约 80%由糖类提供,因 此,它是人类及动物的生命源泉。我国传统膳食习惯是以富含糖类化合物的食物为主食,但 近年来随着动物蛋白质食物产量逐年增加和食品工业的发展,膳食的结构也在逐渐地变化。 一、糖类化合物的种类 糖类化合物的分类常按其组成分为单糖,寡糖和多糖。单糖是一类结构最简单的糖。是 不能再被水解的糖单位。根据其所含原子的数目分为丙糖、丁糖、戊糖和己糖等。根据官能 团的特点又分为醛糖或酮糖。寡糖一般是由 2~10 个分子单糖缩合而成,水解后产生单糖。 多糖是由多个单糖分子缩合而成,多糖中由相同的单糖基组成的称同聚多糖,不相同的单糖 基组成的称杂聚多糖,如按其分子中有无支链,则有直链、支链多糖之分。按其功能不同, 则可分为结构多糖、贮存多糖,抗原多糖等。 二、食品中的糖类化合物 陆地植物和海藻干重的四分之三由糖类化合物构成。谷物、疏菜、果实和可供食用的其 他植物都含有糖类化合物。 大多数植物只含少量蔗糖,大量膳食蔗糖来自经过加工的食品。在加工食品中添加的蔗 糖量一般是比较多的(表 3-1)。蔗糖是从甜菜或甘蔗中分离得到的,果实和蔬菜中只含少量 蔗糖、D-葡萄糖和 D-果糖(表 3-2,表 3-3 和表 3-4)。 谷物只含少量的游离糖,大部分游离糖榆送至种籽中并转变为淀粉,玉米粒含 0.2%~ 0.5%D-葡萄糖、0.1~0.4%D-果糖和 1~2%的蔗糖,小麦粒中这几种糖的含量分别小于 0.1%, 0.1%和 1%
甜玉米具有甜味,是因为采摘时蔗糖尚未全部转变为淀粉。玉米和其他谷物生长期,由 叶片光合作用获得的大部分能量用于合成蔗糖并输送至种籽转变成淀粉。未完全成熟的玉米 含有大量蔗糖,如果采摘适时并迅速煮沸或冷冻,使转化蔗糖的酶被钝化,这样便可保持大 量蔗糖成分,提供可口的膳食。 市场上销售的水果一般是在完全成熟之前采收的,果实有一定硬度利于运输和贮藏。在 贮藏和销售过程中,淀粉在酶的作用下生成蔗糖或其他甜味糖,水果经过这种后熟作用而变 甜变软,这种后熟现象和谷粒、块茎及根中的糖转变为淀粉的过程正好相反。 淀粉是植物中最普通的糖类化合物的能量贮备物,甚至树木的木质部分中也存在淀粉, 而以种籽、根和块茎中最丰富。天然淀粉的结构紧密,在低相对湿度的环境中容易干燥,同 水接触又很快变软,并且能够水解成葡萄糖。 动物产品所含的糖类化合物比其他食品少,肌肉和肝脏中的糖原以及结构与支链淀粉相 似的葡聚糖,都以与淀粉代谢相同的方式进行代谢。 乳糖存在于乳汁中,牛奶中含 4.8%,人乳中为 6.7%,市售液体乳清中为 5%。工业上
甜玉米具有甜味,是因为采摘时蔗糖尚未全部转变为淀粉。玉米和其他谷物生长期,由 叶片光合作用获得的大部分能量用于合成蔗糖并输送至种籽转变成淀粉。未完全成熟的玉米 含有大量蔗糖,如果采摘适时并迅速煮沸或冷冻,使转化蔗糖的酶被钝化,这样便可保持大 量蔗糖成分,提供可口的膳食。 市场上销售的水果一般是在完全成熟之前采收的,果实有一定硬度利于运输和贮藏。在 贮藏和销售过程中,淀粉在酶的作用下生成蔗糖或其他甜味糖,水果经过这种后熟作用而变 甜变软,这种后熟现象和谷粒、块茎及根中的糖转变为淀粉的过程正好相反。 淀粉是植物中最普通的糖类化合物的能量贮备物,甚至树木的木质部分中也存在淀粉, 而以种籽、根和块茎中最丰富。天然淀粉的结构紧密,在低相对湿度的环境中容易干燥,同 水接触又很快变软,并且能够水解成葡萄糖。 动物产品所含的糖类化合物比其他食品少,肌肉和肝脏中的糖原以及结构与支链淀粉相 似的葡聚糖,都以与淀粉代谢相同的方式进行代谢。 乳糖存在于乳汁中,牛奶中含 4.8%,人乳中为 6.7%,市售液体乳清中为 5%。工业上

上采取从乳清中结晶的方法制备乳糖。 第二节糖类化合物的结构 一、单糖 单糖的分子量较小,含有 5 或 6 个碳原于,分子式为 Cn(H2O)m,单糖是 D-甘油醛的衍 生物,如图 3-1 所示。 单糖可以形成缩醛和缩酮,糖分子的羰基可以与糖分子本身的一个醇基反应,形成半缩 醛或半缩酮,分子内的半缩醛或半缩酮,形成五员呋喃糖环或更稳定的六员吡喃糖环。例如, 葡萄糖分子的 C5 羟基和 C1:羟基反应(图 3-2),C5 旋转 180°使氧原子位于环的主平面,而 C6 处在平面的上方,当葡萄糖分子的 C1 成为半缩醛结构中的成分时,它连接 4 个不同的基 团,因而 C1 是手性碳原子,可形成立体构型不同的 α 和β两种异头物。 天然葡萄糖属于 D 异构系列。Α-D 型中异头碳原子 C1 连接的氧原子,与葡萄糖手性碳 原子 C5 的氧原子在分子的同一侧,而β-D 型的 C1 连接的氧原子与 C5 的氧原子处在分子的 异侧。如果用哈沃斯(Haworth)环结构表示,异头碳原子的氧和 C6 在异侧,而β-吡喃葡萄糖 的异头碳原子的氧和哈沃斯环形的羟甲基 C6 在同一侧。 除 C5 外的任何一种手性构型有差别的糖都称为差向异构体,例如,D-甘露糖是 D-葡萄 糖的 C:2 差向异构体,D-半乳糖为 D-葡萄糖的 C4 差向异构体
上采取从乳清中结晶的方法制备乳糖。 第二节糖类化合物的结构 一、单糖 单糖的分子量较小,含有 5 或 6 个碳原于,分子式为 Cn(H2O)m,单糖是 D-甘油醛的衍 生物,如图 3-1 所示。 单糖可以形成缩醛和缩酮,糖分子的羰基可以与糖分子本身的一个醇基反应,形成半缩 醛或半缩酮,分子内的半缩醛或半缩酮,形成五员呋喃糖环或更稳定的六员吡喃糖环。例如, 葡萄糖分子的 C5 羟基和 C1:羟基反应(图 3-2),C5 旋转 180°使氧原子位于环的主平面,而 C6 处在平面的上方,当葡萄糖分子的 C1 成为半缩醛结构中的成分时,它连接 4 个不同的基 团,因而 C1 是手性碳原子,可形成立体构型不同的 α 和β两种异头物。 天然葡萄糖属于 D 异构系列。Α-D 型中异头碳原子 C1 连接的氧原子,与葡萄糖手性碳 原子 C5 的氧原子在分子的同一侧,而β-D 型的 C1 连接的氧原子与 C5 的氧原子处在分子的 异侧。如果用哈沃斯(Haworth)环结构表示,异头碳原子的氧和 C6 在异侧,而β-吡喃葡萄糖 的异头碳原子的氧和哈沃斯环形的羟甲基 C6 在同一侧。 除 C5 外的任何一种手性构型有差别的糖都称为差向异构体,例如,D-甘露糖是 D-葡萄 糖的 C:2 差向异构体,D-半乳糖为 D-葡萄糖的 C4 差向异构体

天然存在的糖环结构实际上并不像哈沃斯表示的投影式平面图,吡喃糖有如下所示的椅 式和船式两种不同构象。 很多己糖以相当刚性的椅式构象存在,只有少数是韧性的船式构象。还有其他几种构象, 例如半倚式和邻位交叉式构象,但都因能量较而不常见。 一般规律是,稳定的环构象是其全部或大多数庞大基团对环轴为平伏键,而环的最小取 代成分(氢)为竖直键。 呋喃糖比吡喃糖稳定性低,可迅速出现所谓信封式和扭曲式的平衡混合物。呋喃糖和吡 喃糖环的构象已经用核磁共振光谱法测定,提供了很多关于单糖在溶液中的构象加识,其结 构模型有助于观察和了解它的三维空间结构。 二、糖苷 如上所述,糖分子的羰基与一个醇基结合生成半缩醛或半缩酮,并在原来羰基位置形成 一个新的手性中心。如果将糖溶解于微酸性乙醇中,半缩醛或半缩酮形式的糖和醇反应生成 缩醛或缩酮。在这种混合缩醛或缩酮产物中,溶剂醇构成分子的一部分,糖本身的醇基是另 一部分,脱水形成的产物称为糖甘。糖苷中的糖部分称为糖基,非糖部分称为配基。 糖苷通常包含一个呋喃糖环或一个吡喃糖环,新形成的手性中心是 α 或β型两种。因此, D-吡喃葡萄糖应看成是 α-D-和β-D 异头体的混合物,形成的糖苷也是。α-D-和β-D-吡喃葡 萄糖苷的混合物。 在酸催化剂作用下生成糖苷的反应是可逆的,若要得到高产率糖昔,在反应过程中必须 除去反应中生成的水。由于吡喃糖苷比呋喃糖苷稳定,所以它是主要的糖苷产物。反应进行
天然存在的糖环结构实际上并不像哈沃斯表示的投影式平面图,吡喃糖有如下所示的椅 式和船式两种不同构象。 很多己糖以相当刚性的椅式构象存在,只有少数是韧性的船式构象。还有其他几种构象, 例如半倚式和邻位交叉式构象,但都因能量较而不常见。 一般规律是,稳定的环构象是其全部或大多数庞大基团对环轴为平伏键,而环的最小取 代成分(氢)为竖直键。 呋喃糖比吡喃糖稳定性低,可迅速出现所谓信封式和扭曲式的平衡混合物。呋喃糖和吡 喃糖环的构象已经用核磁共振光谱法测定,提供了很多关于单糖在溶液中的构象加识,其结 构模型有助于观察和了解它的三维空间结构。 二、糖苷 如上所述,糖分子的羰基与一个醇基结合生成半缩醛或半缩酮,并在原来羰基位置形成 一个新的手性中心。如果将糖溶解于微酸性乙醇中,半缩醛或半缩酮形式的糖和醇反应生成 缩醛或缩酮。在这种混合缩醛或缩酮产物中,溶剂醇构成分子的一部分,糖本身的醇基是另 一部分,脱水形成的产物称为糖甘。糖苷中的糖部分称为糖基,非糖部分称为配基。 糖苷通常包含一个呋喃糖环或一个吡喃糖环,新形成的手性中心是 α 或β型两种。因此, D-吡喃葡萄糖应看成是 α-D-和β-D 异头体的混合物,形成的糖苷也是。α-D-和β-D-吡喃葡 萄糖苷的混合物。 在酸催化剂作用下生成糖苷的反应是可逆的,若要得到高产率糖昔,在反应过程中必须 除去反应中生成的水。由于吡喃糖苷比呋喃糖苷稳定,所以它是主要的糖苷产物。反应进行

一段时间后,几乎不存在呋喃糖苷,只是在糖基化反应刚开始的阶段,呋喃糖苷才占主要部 分。糖基是指除去异头碳上羟基以后剩下的糖残基。 形成糖苷的配基不只是醇基,例如,糖和硫醇 RSH 反应能够得到硫糖苷,与胺(RNH2) 反应生成氨基糖苷。 天然糖苷是糖基从核苷酸衍生物(例如腺苷二磷酸和尿苷二磷酸)中转移至适当的配基 上形成的产物,所生成的糖苷可以是 α 或β型的,这决定于酶的催化专一性。 人类膳食中除低聚糖和多糖外,还有少量糖苷存在。它们的含量虽然不多,但具有重要 的生理效应,例如天然存在的强心苷(毛地黄苷和毛地黄毒苷}、皂角苷(三萜或甾类糖苷), 都是强泡沫形成剂和稳定剂;类黄酮糖苷使食品产生苦味或其他风味和颜色。植物中形成糖 苷有利于那些不易溶解的配基变成可溶于水的物质,这对类黄酮和甾类糖苷特别重要,因为 糖苷形式有利于在水介质中输送。 几种复杂糖苷的甜味很强,例如斯切维苷(stevoside)、奥斯莱丁(osladin)和甘草酸 (glycyrrhizic acid)。但大多数糖昔,特别是当配基部分比甲基大时,则会产生微弱以至极 强的苦味、涩味。醛糖或酮糖均可形成糖苷,例如 D-甘露糖形成缩醛,D-果糖(酮糖)可形成 缩酮。 氧糖苷连接的 O-糖苷在中性和碱性 pH 环境中是稳定的,而在酸性条件下易水解。食 品中(除酸性较强的食品外)大多数糖苷都是稳定的,糖苷在糖苷酶的作用下水解。 对一种新糖苷的鉴定,可以采用化学方法和波谱方法。波谱方法中最适用的是核磁共振 波谱法,用 1~50mg 材料,便可确定异头构型、环构象以及环的大小。 氮糖苷键连接的 N-糖苷不如 O-糖苷稳定,在水中易水解。然而,某些 N-糖苷却十分 稳定,例如 N-葡糖基胺。某些 N-葡糖基嘌呤和嘧啶,特别是次黄嘌呤核苷、黄嘌呤核苷 和鸟嘌呤核苷的 5′-磷酸衍生物,是风味增强剂(见下图)
一段时间后,几乎不存在呋喃糖苷,只是在糖基化反应刚开始的阶段,呋喃糖苷才占主要部 分。糖基是指除去异头碳上羟基以后剩下的糖残基。 形成糖苷的配基不只是醇基,例如,糖和硫醇 RSH 反应能够得到硫糖苷,与胺(RNH2) 反应生成氨基糖苷。 天然糖苷是糖基从核苷酸衍生物(例如腺苷二磷酸和尿苷二磷酸)中转移至适当的配基 上形成的产物,所生成的糖苷可以是 α 或β型的,这决定于酶的催化专一性。 人类膳食中除低聚糖和多糖外,还有少量糖苷存在。它们的含量虽然不多,但具有重要 的生理效应,例如天然存在的强心苷(毛地黄苷和毛地黄毒苷}、皂角苷(三萜或甾类糖苷), 都是强泡沫形成剂和稳定剂;类黄酮糖苷使食品产生苦味或其他风味和颜色。植物中形成糖 苷有利于那些不易溶解的配基变成可溶于水的物质,这对类黄酮和甾类糖苷特别重要,因为 糖苷形式有利于在水介质中输送。 几种复杂糖苷的甜味很强,例如斯切维苷(stevoside)、奥斯莱丁(osladin)和甘草酸 (glycyrrhizic acid)。但大多数糖昔,特别是当配基部分比甲基大时,则会产生微弱以至极 强的苦味、涩味。醛糖或酮糖均可形成糖苷,例如 D-甘露糖形成缩醛,D-果糖(酮糖)可形成 缩酮。 氧糖苷连接的 O-糖苷在中性和碱性 pH 环境中是稳定的,而在酸性条件下易水解。食 品中(除酸性较强的食品外)大多数糖苷都是稳定的,糖苷在糖苷酶的作用下水解。 对一种新糖苷的鉴定,可以采用化学方法和波谱方法。波谱方法中最适用的是核磁共振 波谱法,用 1~50mg 材料,便可确定异头构型、环构象以及环的大小。 氮糖苷键连接的 N-糖苷不如 O-糖苷稳定,在水中易水解。然而,某些 N-糖苷却十分 稳定,例如 N-葡糖基胺。某些 N-葡糖基嘌呤和嘧啶,特别是次黄嘌呤核苷、黄嘌呤核苷 和鸟嘌呤核苷的 5′-磷酸衍生物,是风味增强剂(见下图)