表2-2番茄某些相对性状及F,的自交后代和回交后代之比率 性状 基 杂种第二代观察比例 与隐性亲本回交后代观 因 比例(1:) 显性 隐性 显性 隐性 紫色茎一绿色茎 A-a 3084(3132.7 1093(1044.2 482(505) 526(505) 分 普通叶型一马铃薯叶型 C-c 5473(5510.2 1874(1836.7 694(656. 519(656.5) 5) 5) 6) 总性一锈性 D-d5283(5277) 1753(1759) 560(549) 538(549) 绿色叶一黄色叶 L-15392(5357) 1772(1791) 671(620 569(620) 单花序一多复花序 S-s 752(723.75) 213(241.25) 458(451) 444(451) 果实平滑一多棱沟 F-f 2655(2661) 893(887) 442(434 426(434) 扁平果一椭圆形 0-0 1919(1917) 637639) 521(523) 525(523) 红果肉一黄果肉 R-r 3629(3603.7 1176(1201.2 541(547 553(547)) 5) 5) 黄果肉一透明果皮 4443(4388.21408(1462.7 351(323. 296(323.5) 5) 5) (摘自《番茄育种》,王海廷等编著,1988) 1果实性状的遗传 (1)果形的遗传 长形×圆形、扁形×圆形、卵形×圆形、扁形X卵形的F都是圆形果,李形×梨形的F为李形,: 李形和梨形的分离条例符合3:1:李形×柠檬形的R,为李形,F:分离接近9:6:1的积加效应分离比: 而李形X圆形、柠檬形×扁圆形、牛心形X柠檬形、梨形×柠檬形的,大多为中间型或近圆形,下,出现 多种过渡类型:果顶尖突和果突平滑这一对性状在一些组合中表现为1对基因的分离,在另一些组合中 表现为2对基因间存在关互补效应
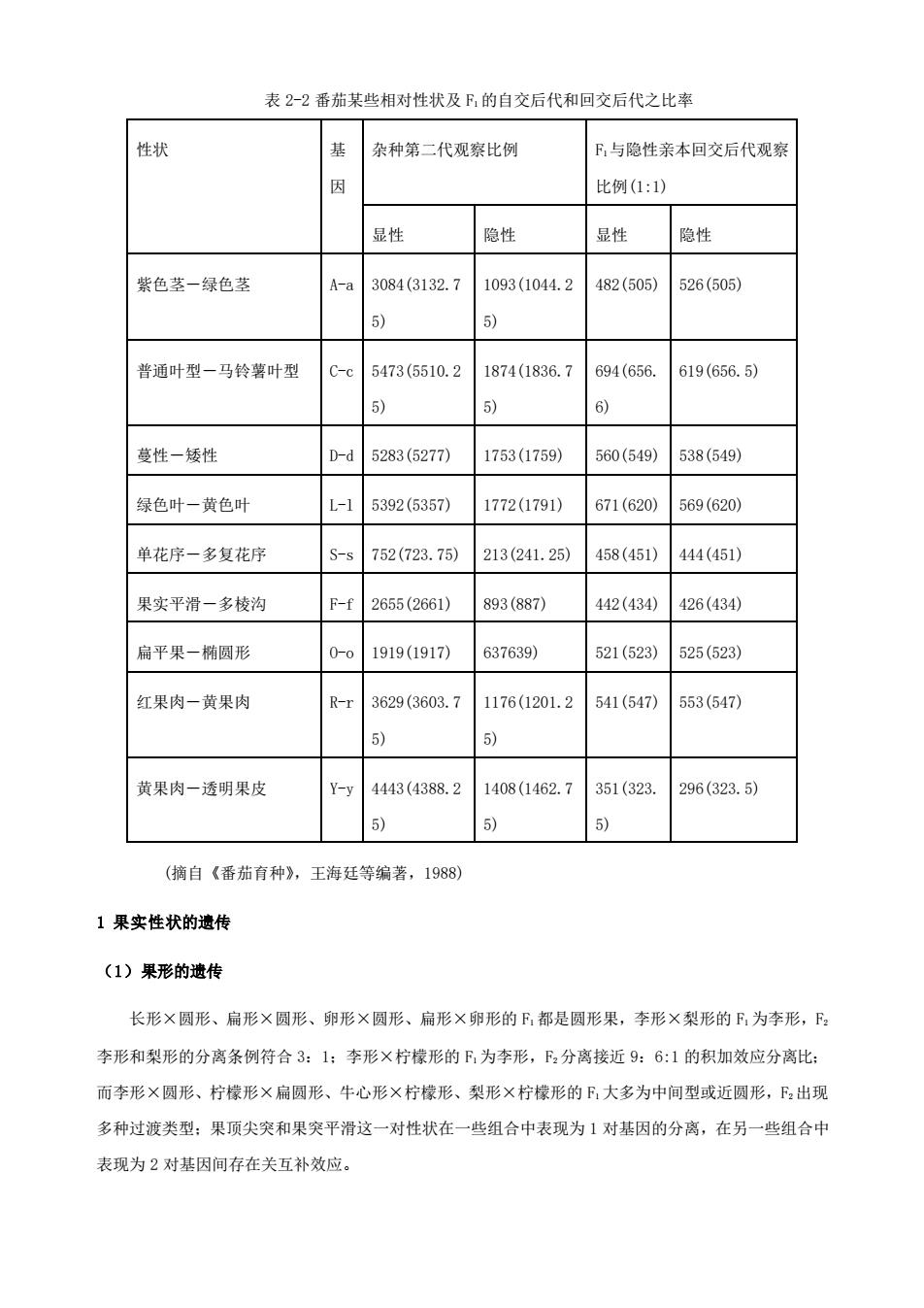
表 2-2 番茄某些相对性状及 F1 的自交后代和回交后代之比率 性状 基 因 杂种第二代观察比例 F1与隐性亲本回交后代观察 比例(1:1) 显性 隐性 显性 隐性 紫色茎-绿色茎 A-a 3084(3132.7 5) 1093(1044.2 5) 482(505) 526(505) 普通叶型-马铃薯叶型 C-c 5473(5510.2 5) 1874(1836.7 5) 694(656. 6) 619(656.5) 蔓性-矮性 D-d 5283(5277) 1753(1759) 560(549) 538(549) 绿色叶-黄色叶 L-l 5392(5357) 1772(1791) 671(620) 569(620) 单花序-多复花序 S-s 752(723.75) 213(241.25) 458(451) 444(451) 果实平滑-多棱沟 F-f 2655(2661) 893(887) 442(434) 426(434) 扁平果-椭圆形 O-o 1919(1917) 637639) 521(523) 525(523) 红果肉-黄果肉 R-r 3629(3603.7 5) 1176(1201.2 5) 541(547) 553(547) 黄果肉-透明果皮 Y-y 4443(4388.2 5) 1408(1462.7 5) 351(323. 5) 296(323.5) (摘自《番茄育种》,王海廷等编著,1988) 1 果实性状的遗传 (1)果形的遗传 长形×圆形、扁形×圆形、卵形×圆形、扁形×卵形的 F1 都是圆形果,李形×梨形的 F1 为李形,F2 李形和梨形的分离条例符合 3:1;李形×柠檬形的 F1 为李形,F2 分离接近 9:6:1 的积加效应分离比; 而李形×圆形、柠檬形×扁圆形、牛心形×柠檬形、梨形×柠檬形的 F1 大多为中间型或近圆形,F2 出现 多种过渡类型;果顶尖突和果突平滑这一对性状在一些组合中表现为 1 对基因的分离,在另一些组合中 表现为 2 对基因间存在关互补效应

(2)果实成熟一致与未熟果肩色的遗传 果实绿色果肩U对果实成熟u为显性:未熟果肩色一致性由2个隐性基因控制,u为一致绿色,g 为一致绿白色。Ayers.J.E.(1996)报道,u u'ug'ug×wwug'ug的R为3:1分离,mug'ug×uuug ug的F为9:3:4分离。另外有1个隐性基因gf控制果实成熟时带绿色果肉。 (3)番茄果色的遗传 用同质的红果番茄与同质的黄果番茄杂交,其F,是红果。果实的颜色取决于果肉色和果皮色2对基 因的配合:R一红色果肉,一黄色果肉,Y一一深黄色果皮,y一一透明果皮,这样,RY表现为红果, Ry表现为粉红果,rY表现橙黄果,y表现为淡黄果。 余诞年指出:果肉颜色分别由2对基因所控制,其中控制红色与黄色的基因系1对等位基因,而橙 色则不是两种颤色的等位基因关系,控制橙色()及对性(T)的基因,对控制红色(R及)的对性基因来 说,为隐性上位关系,B是9:4:3的分离比率。 2番茄雄性不育性的遗传 至1988年,各种类型的雄性不有基因(ms)己先后报道了30个,其中至少有3个功能性不育基 因(s)和6个决定出现无雄蕊的基因sl和sTs。番茄雄性不有的表现主要可分为:功能不有(型)、雄 蕊退化(6型)、花粉不有、部位不有()四种类型。 番茄的雄性不育属于“核质型”(Genetietype))。Larson等(I954)认为,番茄的雄性不育系由 1对简单的独立核基因(sms)支配,余诞年的研究指出:雄蕊退化×功能不有,其F,为正常花,F:分离 为正常花(44)、雄蕊退化(10)、功能不有(15)、雄蕊退化兼功能不育(⑤):功能不有×长花柱,F,为正 常花,B:分离为正常花(45)、长花柱(12)、功能不有(12、长花柱功能不有(8)、均符合3:1或9:3: 3:1的孟德尔遗传比率。 3番茄几种病害的抗性遗传 ()烟草花叶病毒病(TW) 己知可供番茄有种使用的抗TW基因有3个,即T-1、T-2Tm-2。单显性基因Tm-1可以减轻对 TMW侵染的敏感性:显性基因T一2具有比Tl基因强的抗性,但与隐性黄基因v连锁,在纯合情况下 引起植株矮化和黄化:单显性基因Tm一2(即Tm-2)与Tm-2基因呈紧密连锁但又不与v基因连锁,Tm-2 和Tr一2对TW病毒株系(Strain)的反映有明显的差异,它们各自可以成为纯合系统,也可以得到抗病 性更广泛的Tm一2/Tm2杂合系统。用不同基因型抗性的亲本进行杂交,其后代(,)的抗性遗传表现如 下
(2)果实成熟一致与未熟果肩色的遗传 果实绿色果肩 U 对果实成熟 u 为显性;未熟果肩色一致性由 2 个隐性基因控制,u 为一致绿色,ug 为一致绿白色。Ayers.J.E.(1996)报道,u + u + ug + ug +×uuug + ug +的 F2 为 3:1 分离, uuug + ug +×u + u + ug + ug +的 F2 为 9:3:4 分离。另外有 1 个隐性基因 gf 控制果实成熟时带绿色果肉。 (3)番茄果色的遗传 用同质的红果番茄与同质的黄果番茄杂交,其 F1 是红果。果实的颜色取决于果肉色和果皮色 2 对基 因的配合;R-红色果肉,r-黄色果肉,Y-深黄色果皮,y-透明果皮,这样,RY 表现为红果, Ry 表现为粉红果,rY 表现橙黄果,ry 表现为淡黄果。 余诞年指出:果肉颜色分别由 2 对基因所控制,其中控制红色与黄色的基因系 1 对等位基因,而橙 色则不是两种颜色的等位基因关系,控制橙色(t)及对性(T)的基因,对控制红色(R 及 r)的对性基因来 说,为隐性上位关系,F2是 9:4:3 的分离比率。 2 番茄雄性不育性的遗传 至 1988 年,各种类型的雄性不育基因(ms)已先后报道了 30 个,其中至少有 3 个功能性不育基 因(ps)和 6 个决定出现无雄蕊的基因 sl 和 sTs。番茄雄性不育的表现主要可分为:功能不育(J 型)、雄 蕊退化(S 型)、花粉不育、部位不育(L)四种类型。 番茄的雄性不育属于“核质型”(Genetietype)。Larson 等(1954)认为,番茄的雄性不育系由 1 对简单的独立核基因(msms)支配,余诞年的研究指出:雄蕊退化×功能不育,其 F1 为正常花,F2 分离 为正常花(44)、雄蕊退化(10)、功能不育(15)、雄蕊退化兼功能不育(5);功能不育×长花柱,F1 为正 常花,F2 分离为正常花(45)、长花柱(12)、功能不育(12)、长花柱功能不育(8)、均符合 3:1 或 9:3: 3:1 的孟德尔遗传比率。 3 番茄几种病害的抗性遗传 (1) 烟草花叶病毒病(TMV) 已知可供番茄育种使用的抗 TMV 基因有 3 个,即 Tm-1、 Tm-2 Tm-2 a。单显性基因 Tm-1 可以减轻对 TMV 侵染的敏感性;显性基因 Tm-2 具有比 Tm-1 基因强的抗性,但与隐性黄基因 nv 连锁,在纯合情况下 引起植株矮化和黄化;单显性基因 Tm-2 a (即 Tm-2 2 )与 Tm-2 基因呈紧密连锁但又不与 nv 基因连锁,Tm-2 a 和 Tm-2 对 TMV 病毒株系(Strain)的反映有明显的差异,它们各自可以成为纯合系统,也可以得到抗病 性更广泛的 Tm-2/ Tm-2 a 杂合系统。用不同基因型抗性的亲本进行杂交,其后代(F1)的抗性遗传表现如 下

(2)番茄青枯病(Pseudomona.Solanacearum E.R.Smith) 番茄青枯病的抗性遗传规律日前仍不十分清楚。有人认为抗性是隐性遗传,也有人认为抗病对感病 为不完全显性,杂交一代的抗性表现主要依靠双亲提供各自一半的抗性遗传成分,单靠显性成分在F 代的优势是不够的,控制青枯病抗性的基因数目有认为是多数,亦有认为是单个显性基因。由于番茄对 青枯病的抗性遗传比较复杂 不同研究者用不同的试验材料进行研究,所得结论不尽相同。李海涛等以感病材料98-1,L 402本自交系、高抗青枯病自交系LS89及感、抗材料配制杂交组合和回交世代为试材,进行番茄抗青 枯病遗传机制研究。结果表明:抗病基因对感病基因表现为部分显性:控制LS89番茄抗青枯病自交系 的遗传是一个基因,或一个基因起主导作用:番茄抗青枯病抗性遗传受核基因控制,细胞质基因对其影 响不大。乐素菊等以不同抗青枯病水平的番茄品种(品系)进行5×5双列杂交的遗传分析表明:番茄对 青枯病的抗性至少由3对基因控制:在抗性遗传效应中,加性成分占主导地位:广义遗传力为97第, 狭义遗传力为69%:抗病对感病为不完全显性。邓铭光等的研究认为苗期抗性存在加性和显性基因效应 以加性效应为主:抗性的狭义遗传力为63.36%:但抗病对感病为隐性。在育种中不能期望下1有抗性 杂种优势,只有通过杂交选择积累抗性基因,培有抗性更高的有种系,然后再行配组。 (3)番茄叶幂病(Cladosporum fulvum Cooke) 经研究确认,番茄叶霉病抗性受显性基因控制,抗性为显性,大多表现为质量性状遗传。1980听 Kanwar.J.S.等把所有24个抗叶霉病基因全部定位于番茄12条染色体上。 (④)番茄枯菱病(Fusarium oxysporum f.lycopersici Sacc) 目前已发现生理小种1、2、3、及3个显性抗病基因,即基因I、【-2、1-3。大多数认为,番茄 枯菱病的抗性由一主要显性基因控制,另有修饰基因:也有人认为抗性基因为不完全显性。 (5)番茄早疫病[Alternaria solani(E.etM)Jones et Gront](Mecrosporium solani Ell.et Mart.) 抗早疫病遗传性有人认为受不完全单显性基因支配,并命名为Ad:亦有认为,此病为单基因控制, 抗性为隐性,或感病为不完全显性:Barksdale T.H和Stoner A.K(1977)指出,抗病的77B含有2个或 2个以上的叶部抗早疫病的隐性遗传基因,认为实际抗性遗传较为复杂。 ()番茄晚疫病[Phytophthora infestans(mont.)Bary] 番茄抗晚疫病性状由两类不同的基因控制,一种受单显性基因h控制,对生理小种0兔疫:另 种受多基因控制,与多种因素有关,属数量遗传,对生理小种0和生理小种1都有效.西弗吉尼亚63(s irginia3)同时存在若单显性基因抗性和多基因抗性。英国品种原子tom)带有2个不完全显性的水
(2) 番茄青枯病(Pseudomonas Solanacearum E.F.Smith) 番茄青枯病的抗性遗传规律目前仍不十分清楚。有人认为抗性是隐性遗传,也有人认为抗病对感病 为不完全显性,杂交一代的抗性表现主要依靠双亲提供各自一半的抗性遗传成分,单靠显性成分在 F1 代的优势是不够的,控制青枯病抗性的基因数目有认为是多数,亦有认为是单个显性基因。由于番茄对 青枯病的抗性遗传比较复杂, 不同研究者用不同的试验材料进行研究,所得结论不尽相同。李海涛等以感病材料 98TL - 1 ,L - 402 本自交系、高抗青枯病自交系 LS89 及感、抗材料配制杂交组合和回交世代为试材,进行番茄抗青 枯病遗传机制研究。结果表明:抗病基因对感病基因表现为部分显性;控制 LS89 番茄抗青枯病自交系 的遗传是一个基因,或一个基因起主导作用;番茄抗青枯病抗性遗传受核基因控制,细胞质基因对其影 响不大。乐素菊等以不同抗青枯病水平的番茄品种(品系) 进行 5 ×5 双列杂交的遗传分析表明:番茄对 青枯病的抗性至少由 3 对基因控制;在抗性遗传效应中,加性成分占主导地位;广义遗传力为 97 % , 狭义遗传力为 69 %;抗病对感病为不完全显性。邓铭光等的研究认为苗期抗性存在加性和显性基因效应, 以加性效应为主;抗性的狭义遗传力为 63.36 %;但抗病对感病为隐性。在育种中不能期望 F1 有抗性 杂种优势,只有通过杂交选择积累抗性基因,培育抗性更高的育种系,然后再行配组。 (3) 番茄叶霉病(Cladosporum fulvum Cooke) 经研究确认,番茄叶霉病抗性受显性基因控制,抗性为显性,大多表现为质量性状遗传。1980 昕 Kanwar.J.S.等把所有 24 个抗叶霉病基因全部定位于番茄 12 条染色体上。 (4) 番茄枯萎病(Fusarium oxysporum f.lycopersici Sacc) 目前已发现生理小种 1、2、3、及 3 个显性抗病基因,即基因Ⅰ、Ⅰ-2、Ⅰ-3。大多数认为,番茄 枯萎病的抗性由一主要显性基因控制,另有修饰基因;也有人认为抗性基因为不完全显性。 (5) 番茄早疫病[Alternaria solani(E.et M.)Jones et Gront](Mecrosporium solani Ell.et Mart.) 抗早疫病遗传性有人认为受不完全单显性基因支配,并命名为 Ad;亦有认为,此病为单基因控制, 抗性为隐性,或感病为不完全显性;Barksdale T.H 和 Stoner A.K(1977)指出,抗病的 77B2 含有 2 个或 2 个以上的叶部抗早疫病的隐性遗传基因,认为实际抗性遗传较为复杂。 (6) 番茄晚疫病[Phytophthora infestans (mont.) Bary] 番茄抗晚疫病性状由两类不同的基因控制,一种受单显性基因 ph 控制,对生理小种 0 免疫;另一 种受多基因控制,与多种因素有关,属数量遗传,对生理小种 0 和生理小种 1 都有效。西弗吉尼亚 63(West Virginia 63)同时存在着单显性基因抗性和多基因抗性。英国品种原子(Atom)带有 2 个不完全显性的水

平抗性的主效基因phf-1、phf-2,对0系和1系都有抗性,此外还有垂直抗病性基因ph-1、ph-2以及 多基因控制的抗性, (7)番茄根痛线虫病(Meloidogyne ssp.) 番茄对线虫的抗性是由主效显性基因Mi决定的,能抗M.javanica、M arenaria三种线虫。抗X不 抗的F,全为抗病,F,与不抗病的亲本回交后有50%为抗性杂合体。1974年,Rick和Fobes发现,抗线虫 品种都带有酸性磷酸酶的一个变体APS-1',认为以PS-'基因的存在与否为标准进行选择是很有效的。 (⑧)番茄叶斑枯病(Septoria lycopersici Speg) 对此病有高抗性的一些品种(如Targinnie Red),其抗性由显性基因控制,源自毛番茄PI3833的抗性 为隐性。 (9炭疽病[Colletotrichum phomoides(Sace.)Chester.] 番茄对炭疽病的抗性由多基因控制,抗性为部分显性。 (10)番茄黄菱病 番茄抗黄萎病生理小种1的强弱或取决于一个多基因系统或取决于一个单个控制基因Ve,V是 由一个名为peruvian的野生番茄衍生而来(Schaibil,l.l951)。利用LP进行Ve的制图,认为 Ve基因位于番茄的第9染色体的短臂上,单一位点紧密地与RFLP基因标记GP39相连,这就排除了 黄菱病抗性被一个多位点系统控制的可能性,这一点也体现在番茄的Bm抗性和TYLCV耐性。 4番茄抗逆性遗传 研究结果表明:在高温下番茄的花数是受隐性基因控制的,其遗传务较高:坐果率则受加性基因 控制,遗传力中等:结籽率无等位基因互作,显性基因作用超过加性效应,遗传力较低;高温造成的长 柱头现象是受部分显性基因和加性效应共同控制的,其遗传力较高。而在低温下番茄花粉萌发是受单基 因控制的。高海拔种多毛番茄、智利番茄、类番茄比低海拔的栽培品种具有更高的抗寒性。 (1)番茄抗冷性的遗传 番茄抗冷性是复杂的数量性状,受微效多基因控制,其性状表现易受环境条件的影响。下oolad等 对番茄种子发芽期抗冷性的遗传进行了研究,他们以番茄抗冷品系(PI120256)和冷敏感品系(UCT5) 的种子为试材对发芽期耐冷性研究的结果表明在低温胁迫条件下,种子发芽期抗冷性的遗传变异主要 取决于加性遗传效应,而显性效应和上位性效应不明显。进一步研究表明在杂种2代对种子发芽期的 耐冷性进行选择有明显效果,在3代经选择的材料与未经选择的对照相比抗冷性有明显增强。对番茄
平抗性的主效基因 phf-1、phf-2,对 0 系和 1 系都有抗性,此外还有垂直抗病性基因 ph-1、ph-2 以及 多基因控制的抗性。 (7) 番茄根瘤线虫病(Meloidogyne ssp.) 番茄对线虫的抗性是由主效显性基因 Mi 决定的,能抗 M.javanica、M.arenaria 三种线虫。抗×不 抗的 F1 全为抗病,F1 与不抗病的亲本回交后有 50%为抗性杂合体。1974 年,Rick 和 Fobes 发现,抗线虫 品种都带有酸性磷酸酶的一个变体 APS-1 1,认为以 APS-1 1 基因的存在与否为标准进行选择是很有效的。 (8) 番茄叶斑枯病(Septoria lycopersici Speg) 对此病有高抗性的一些品种(如 Targinnie Red)其抗性由显性基因控制,源自毛番茄 PI3833 的抗性 为隐性。 (9) 炭疽病[Colletotrichum phomoides (Sace.)Chester.] 番茄对炭疽病的抗性由多基因控制,抗性为部分显性。 (10)番茄黄萎病 番茄抗黄萎病生理小种 1 的强弱或取决于一个多基因系统或取决于一个单个控制基因 Ve ,Ve 是 由一个名为 peruvian 的野生番茄衍生而来( Schaibil. l.1951) 。利用 RFLP 进行 Ve 的制图,认为 Ve 基因位于番茄的第 9 染色体的短臂上,单一位点紧密地与 RFL P 基因标记 GP39 相连,这就排除了 黄萎病抗性被一个多位点系统控制的可能性,这一点也体现在番茄的 BW 抗性和 TYLCV 耐性。 4 番茄抗逆性遗传 研究结果表明:在高温下番茄的花数是受隐性基因控制的,其遗传务较高;坐果率则受加性基因 控制,遗传力中等;结籽率无等位基因互作,显性基因作用超过加性效应,遗传力较低;高温造成的长 柱头现象是受部分显性基因和加性效应共同控制的,其遗传力较高。而在低温下番茄花粉萌发是受单基 因控制的。高海拔种多毛番茄、智利番茄、类番茄比低海拔的栽培品种具有更高的抗寒性。 (1)番茄抗冷性的遗传 番茄抗冷性是复杂的数量性状, 受微效多基因控制, 其性状表现易受环境条件的影响。Foolad 等 对番茄种子发芽期抗冷性的遗传进行了研究, 他们以番茄抗冷品系(PI120256) 和冷敏感品系〔UCT5) 的种子为试材对发芽期耐冷性研究的结果表明在低温胁迫条件下, 种子发芽期抗冷性的遗传变异主要 取决于加性遗传效应, 而显性效应和上位性效应不明显。进一步研究表明在杂种 F2 代对种子发芽期的 耐冷性进行选择有明显效果, 在 F3 代经选择的材料与未经选择的对照相比抗冷性有明显增强。对番茄

营养生长期植株抗冷性的研究结果也表明抗冷性主要是由加性效应决定的。KozLova等认为番茄的抗冷 性受到核基因和细胞质基因控制,也与核质基因的互作有关。他们在孢子体和雄性配子体水平上对抗 性进行选择,获得了有商品价值的F1杂种。林多等对低温下番茄幼苗生长遗传规律进行了研究。选用 4份耐低温能力不同的番茄品种(系)为亲本,采用完全双列杂交设计模式,调查适温、低温条件下 的幼苗生长速率,应用Hayman和Griffing的数量分析方法进行遗传分析。结果表明低温下番茄幼苗 生长的遗传为不完全显性,回交效应显著,正反交差异不显著,呈现核遗传,细胞质作用不显著, 符合“加性-显性”遗传模型,且加性效应更为重要,亲代与子代间存在极显著的正相关。 近年来植物分子标记技术迅速发展,为数量性状基因作图创造了条件。目前已用FLP(Rest riction Fragment Length Polymorphis,限制性片段长度多态性)分子标记构建了番茄的基因连锁 图,有3个与番茄种子发芽期抗冷性相关的基因定位在】号染色体上,另有2个抗冷性基因分别定 位于4号和8号染色体上。近年来的研究还发现番茄种子发芽期的抗冷性主要是由少数几个主要QTL (Quantitative Trait Loci,数量性状位点)决定的。因此,采用分子标记辅助选择对番茄抗冷性 进行遗传改良可能是有效的途径。 (2)番茄耐热性遗传规律研究动态 关于耐热性遗传规律的研究较少,有关蔬菜耐热性的研究大多集中在大白菜、萝卜等上。研究抗热与热 傲感性的番茄杂交双亲地F1,2和C1表明,正常花的比例和结实率呈部分显性,结实性具有一定的 杂种优势。热胁迫下,着花数和结果性在遗传上似乎是独立的。双列杂交分析初步表明,番茄坐果的耐 热表现为加性遗传。用6个花器官对热敏反应不同的番茄品种配制双列杂交试验表明,许多品种的柱头 伸出程度、同化物向果实的分配、散粉和花粉活力存在差异。这些性状的组合导致热胁迫下结实率的巨 大差异,并使每个性状获得一个抗性遗传途径。在正常温度和高条件下,花与高遗传力隐性基因有关, 加性遗传方差参与控制结实率,它具有中等遗传力,上位基因互作参与控制结籽,其显性部分比加性 部分控制更大。高温下,柱头伸出多受遗传力高的部分显性基因控制,对特殊配合力进行选择有助于抗 热性与其他优良农艺性状的重组。研究表明耐热性由数量遗传所控制。许多研究认为正常温度下柱头位 置遗传性是一个分离性状。用测量柱头和雄蕊孔间的距离来说明柱头伸出的程度,发现这个性状由单基 因控制。高温下,柱头伸长的遗传研究发现,这个性状的广义遗传力0.43,结籽是雌雄配子可育的指标 这一性状存在非等位基因互作。目前己知的单性结实基因有5个:pat,阳t2,p阳t3,阳t4,pat5。可见 番茄耐热性遗传规律是复杂多样的。植物中除环境与基因型的互作外,尚存在明显的核质互作现象,并 进一步推断,耐热性基因与其他生长性状间存在者复杂的连锁关系,故影响性状的表现。番茄耐热性的 遗传规律是目前番茄耐热性研究领域中比较薄弱的环节,而目前仍是以传统育种为主要方式来对作物进 行改良。因此,这方面的研究对杂交育种中亲本的正确选择、选配具有重要的指导意义
营养生长期植株抗冷性的研究结果也表明抗冷性主要是由加性效应决定的。KozLova 等认为番茄的抗冷 性受到核基因和细胞质基因控制, 也与核质基因的互作有关。他们在孢子体和雄性配子体水平上对抗 性进行选择, 获得了有商品价值的 F1 杂种。林多等对低温下番茄幼苗生长遗传规律进行了研究。选用 4 份耐低温能力不同的番茄品种(系) 为亲本, 采用完全双列杂交设计模式, 调查适温、低温条件下 的幼苗生长速率, 应用 Hayman 和 Griffing 的数量分析方法进行遗传分析。结果表明低温下番茄幼苗 生长的遗传为不完全显性, 回交效应显著, 正反交差异不显著, 呈现核遗传, 细胞质作用不显著, 符合“加性- 显性”遗传模型, 且加性效应更为重要, 亲代与子代间存在极显著的正相关。 近年来植物分子标记技术迅速发展, 为数量性状基因作图创造了条件。目前已用 RFLP (Rest riction Fragment Length Polymorphism , 限制性片段长度多态性) 分子标记构建了番茄的基因连锁 图, 有 3 个与番茄种子发芽期抗冷性相关的基因定位在 l 号染色体上, 另有 2 个抗冷性基因分别定 位于 4 号和 8 号染色体上。近年来的研究还发现番茄种子发芽期的抗冷性主要是由少数几个主要 QTL (Quantitative Trait Loci , 数量性状位点) 决定的。因此, 采用分子标记辅助选择对番茄抗冷性 进行遗传改良可能是有效的途径。 (2)番茄耐热性遗传规律研究动态 关于耐热性遗传规律的研究较少,有关蔬菜耐热性的研究大多集中在大白菜、萝卜等上。研究抗热与热 敏感性的番茄杂交双亲地 F1,F2 和 BC1 表明,正常花的比例和结实率呈部分显性,结实性具有一定的 杂种优势。热胁迫下,着花数和结果性在遗传上似乎是独立的。双列杂交分析初步表明,番茄坐果的耐 热表现为加性遗传。用 6 个花器官对热敏反应不同的番茄品种配制双列杂交试验表明,许多品种的柱头 伸出程度、同化物向果实的分配、散粉和花粉活力存在差异。这些性状的组合导致热胁迫下结实率的巨 大差异,并使每个性状获得一个抗性遗传途径。在正常温度和高条件下,花与高遗传力隐性基因有关, 加性遗传方差参与控制结实率,它具有中等遗传力,上位基因互作参与控制结籽,其显性部分比加性 部分控制更大。高温下,柱头伸出多受遗传力高的部分显性基因控制,对特殊配合力进行选择有助于抗 热性与其他优良农艺性状的重组。研究表明耐热性由数量遗传所控制。许多研究认为正常温度下柱头位 置遗传性是一个分离性状。用测量柱头和雄蕊孔间的距离来说明柱头伸出的程度,发现这个性状由单基 因控制。高温下,柱头伸长的遗传研究发现,这个性状的广义遗传力0.43,结籽是雌雄配子可育的指标, 这一性状存在非等位基因互作。目前已知的单性结实基因有5个:pat,pat2,pat3,pat4,pat5。可见 番茄耐热性遗传规律是复杂多样的。植物中除环境与基因型的互作外,尚存在明显的核质互作现象,并 进一步推断,耐热性基因与其他生长性状间存在着复杂的连锁关系,故影响性状的表现。番茄耐热性的 遗传规律是目前番茄耐热性研究领域中比较薄弱的环节,而目前仍是以传统育种为主要方式来对作物进 行改良。因此,这方面的研究对杂交育种中亲本的正确选择、选配具有重要的指导意义