
第十三章 代谢调节 新陈代谢是指生物体内的一切分解和合成的作用,它是生物最基本的特征之一。新陈 代谢有其自身的特点。首先,新陈代谢过程是由一系列多步骤的中间反应组成的。这些反 应互相协调和制约,有条不紊、有序地进行。第二,代谢反应是在比较温和的条件下进行 的,绝大多数都由酶催化,表现出灵活的自我调节。代谢物通过影响酶的活性和酶量的变 化,改变合成和分解反应的速度,这是一切调节的基础。多步骤反应受多个酶的调节,因 而有较大的可变性和适应性,而且能最有效地利用能量。第三,代谢调节是生物在长期演 化过程中,为适应环境需要而形成的。新陈代谢既是一个互相联系、错综复杂的变化过程, 又是一个通过机体自身调节使之保持相对平衡的稳定过程。 代谢周节是指细胞内的代谢速度按照生物的需要而记改变的一种生理作用。进化愈高的 生物,其代谢调节的机构就愈复杂,随着生物由单细胞进化为多细胞,除了在细胞和分子 水平的调节外,还有更高层次的激素水平(组织和器官)和整体水平神经和维管束系统)的调 节。本章主要介绍生物体内各类物质代谢的相互联系,以及细胞和分子水平的代谢调节。 细胞和分子水平的调节可有两种方式:一种是酶量的调节,为缓慢调节类型。它是通 过改变酶分子合成或降解的速度来改变细胞内酶的含量。另一种是酶活性的调节,属于快 速调节类型。它是通过酶分子结构的改变来实现对酶促反应速度的调节。 酶在细胞内有一定的布局,互相有关的酶往往组成一个多酶系统而分布于胞内特定部 位,这些酶互相靠近、容易接触,使反应迅速进行:而其他酶系则分布在不同部位,各不 相扰。这样的分隔分布为细胞水平的精确调节创造了有利条件,以保证代谢顺利进行 生物机体的代谢是和机体的内外环境分不开的,生物具有适应环境的能力,当内外条 件改变时,生物机体能调整和改变其体内的代谢过程,建立新的代谢平衡,以适应变化了 的环境,因此代谢平衡是动态的、相对的。生物体生长发育的全过程就是一个不断地进行 代谢调节的过程。 第一节代谢途径的相互联系 生物界,包括人类、动物、植物和微生物,尽管其结构特征和生活方式多种多样,千 变万化。然而,它们却有着共同的最基本的新陈代谢过程,这表明地球上的生物有统一的 起源。 所有细胞都是由多糖、脂类复合物、蛋白质和核酸四类生物大分子,为数有限的生物 321
321 第十三章 代谢调节 新陈代谢是指生物体内的一切分解和合成的作用,它是生物最基本的特征之一。新陈 代谢有其自身的特点。首先,新陈代谢过程是由一系列多步骤的中间反应组成的。这些反 应互相协调和制约,有条不紊、有序地进行。第二,代谢反应是在比较温和的条件下进行 的,绝大多数都由酶催化,表现出灵活的自我调节。代谢物通过影响酶的活性和酶量的变 化,改变合成和分解反应的速度,这是一切调节的基础。多步骤反应受多个酶的调节,因 而有较大的可变性和适应性,而且能最有效地利用能量。第三,代谢调节是生物在长期演 化过程中,为适应环境需要而形成的。新陈代谢既是一个互相联系、错综复杂的变化过程, 又是一个通过机体自身调节使之保持相对平衡的稳定过程。 代谢调节是指细胞内的代谢速度按照生物的需要而改变的一种生理作用。进化愈高的 生物,其代谢调节的机构就愈复杂,随着生物由单细胞进化为多细胞,除了在细胞和分子 水平的调节外,还有更高层次的激素水平(组织和器官)和整体水平(神经和维管束系统)的调 节。本章主要介绍生物体内各类物质代谢的相互联系,以及细胞和分子水平的代谢调节。 细胞和分子水平的调节可有两种方式:一种是酶量的调节,为缓慢调节类型。它是通 过改变酶分子合成或降解的速度来改变细胞内酶的含量。另一种是酶活性的调节,属于快 速调节类型。它是通过酶分子结构的改变来实现对酶促反应速度的调节。 酶在细胞内有一定的布局,互相有关的酶往往组成一个多酶系统而分布于胞内特定部 位,这些酶互相靠近、容易接触,使反应迅速进行;而其他酶系则分布在不同部位,各不 相扰。这样的分隔分布为细胞水平的精确调节创造了有利条件,以保证代谢顺利进行。 生物机体的代谢是和机体的内外环境分不开的,生物具有适应环境的能力,当内外条 件改变时,生物机体能调整和改变其体内的代谢过程,建立新的代谢平衡,以适应变化了 的环境,因此代谢平衡是动态的、相对的。生物体生长发育的全过程就是一个不断地进行 代谢调节的过程。 第一节 代谢途径的相互联系 生物界,包括人类、动物、植物和微生物,尽管其结构特征和生活方式多种多样,千 变万化。然而,它们却有着共同的最基本的新陈代谢过程,这表明地球上的生物有统一的 起源。 所有细胞都是由多糖、脂类复合物、蛋白质和核酸四类生物大分子,为数有限的生物
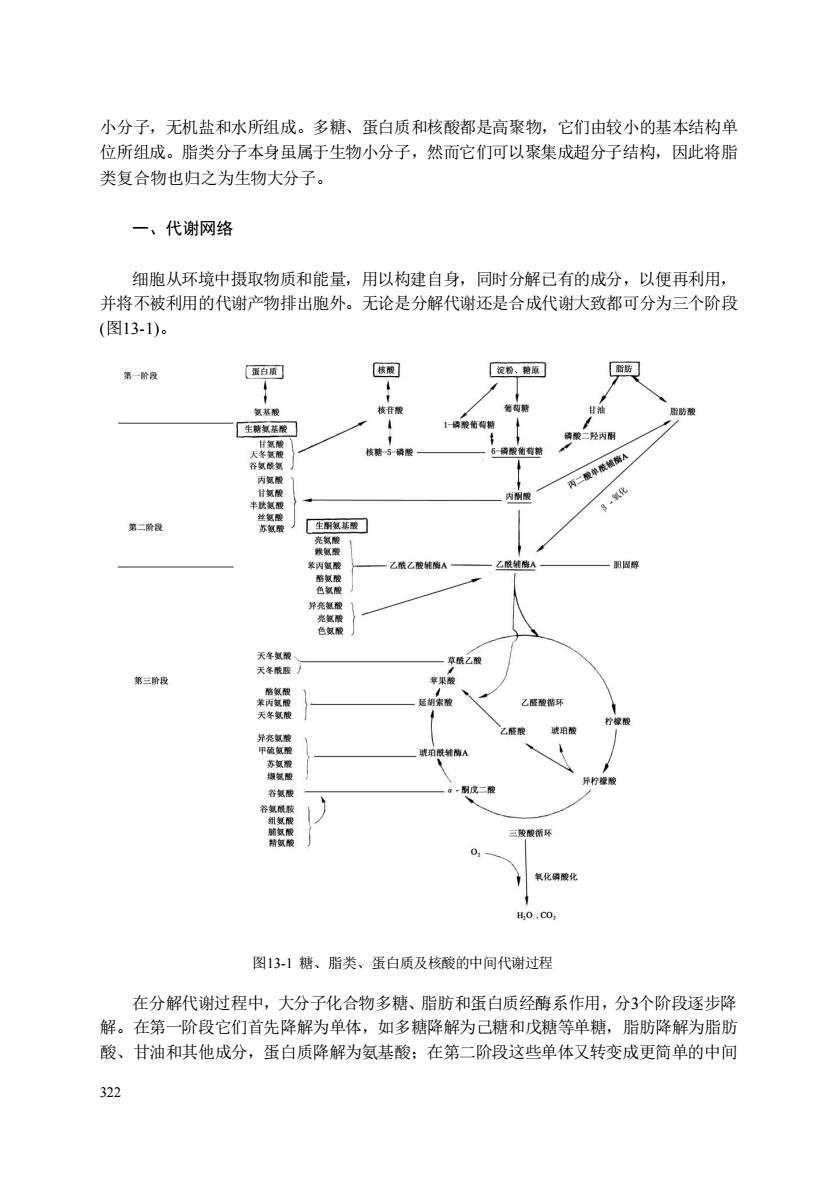
小分子,无机盐和水所组成。多糖、蛋白质和核酸都是高聚物,它们由较小的基本结构单 位所组成。脂类分子本身虽属于生物小分子,然而它们可以聚集成超分子结构,因此将脂 类复合物也归之为生物大分子。 一、代谢网络 细胞从环境中摄取物质和能量,用以构建自身,同时分解己有的成分,以便再利用, 并将不被利用的代谢产物排出胞外。无论是分解代谢还是合成代谢大致都可分为三个阶段 (图13-1)。 第阶 蛋 第二 天货 第三险 智 图131糖、脂类、蛋白质及核酸的中间代谢过程 在分解代谢过程中,大分子化合物多糖、脂肪和蛋白质经酶系作用,分3个阶段逐步降 解。在第一阶段它们首先降解为单体,如多糖降解为己糖和戊糖等单糖,脂肪降解为脂肪 酸、甘油和其他成分,蛋白质降解为氨基酸:在第二阶段这些单体又转变成更简单的中间
322 小分子,无机盐和水所组成。多糖、蛋白质和核酸都是高聚物,它们由较小的基本结构单 位所组成。脂类分子本身虽属于生物小分子,然而它们可以聚集成超分子结构,因此将脂 类复合物也归之为生物大分子。 一、代谢网络 细胞从环境中摄取物质和能量,用以构建自身,同时分解已有的成分,以便再利用, 并将不被利用的代谢产物排出胞外。无论是分解代谢还是合成代谢大致都可分为三个阶段 (图13-1)。 图13-1 糖、脂类、蛋白质及核酸的中间代谢过程 在分解代谢过程中,大分子化合物多糖、脂肪和蛋白质经酶系作用,分3个阶段逐步降 解。在第一阶段它们首先降解为单体,如多糖降解为己糖和戊糖等单糖,脂肪降解为脂肪 酸、甘油和其他成分,蛋白质降解为氨基酸;在第二阶段这些单体又转变成更简单的中间

代谢物,如己糖、戊糖、生糖氨基酸和甘油降解为丙酮酸然后生成乙酰辅酶A,同样脂肪 酸和生酮氨基酸也降解为乙酰辅酶A和其他几个末端产物:最后阶段是乙酰辅酶A和其他产 物氧化成水和CO2。 合成代谢则经历反向的三个阶段,首先是以分解代谢第三阶段中生成的(或从环境中 摄入的)小分子作为起始原料合成简单有机物。不同的生物所利用的起始原料很不相同, 基本上分为二类,一类以二氧化碳为同化起始物合成各种有机物质(称为自养生物),不 需要由外界提供有机的碳化合物,如高等绿色植物、蓝绿藻、光合细菌、硝酸和亚硝酸细 菌等。另一类只能利用有机碳化物为起始原料(称为异养生物),如所有较高等的动物、 无光合作用的植物细胞、大多数微生物等。第二阶段是简单的有机物进一步合成构造大分 子所需要的单体。如ā酮酸与氨基供体反应生成ā氨基酸,磷酸丙糖生成磷酸己糖和磷 酸戊糖。丙酮酸经糖原异生作用生成的单酰乙酰辅酶A可缩合生成脂肪酸,磷酸二羟丙酮 可还原为甘油。第三阶段是由单体合成大分子化合物,如ā氨基酸结合成多肽:磷酸己糖 合成多糖:磷酸戊糖先形成核苷酸,然后合成核酸:甘油和脂肪酸合成脂肪。 分解代谢从各种不同的多糖、脂肪、蛋白质开始逐步形成共同的中间代谢物到第三阶 段则集中到 一个共同的代谢途径三羧酸循环中。当然,在复杂的代谢中第二阶段的氨基酸 甘油、单糖等单体并不是都要经过三羧酸循环,也可以直接作为合成大分子的中间物。合 成代谢则与之相反, 一开始是共同的少数起始原料,然后逐步分化,经过第二阶段到第 阶段产生了许多分枝途径,合成了许多不同类型的生物大分子。生物大分子具有高度的特 异性,正是它们决定了生物物种和个体之间的差异。生物机体将各类物质分别纳入各自的 共同代谢途径,以少数种类的反应,如氧化还原、基团转移、水解脱水、裂解合成、异构 反应等,转化成种类繁多的分子。不同的代谢途径可通过交叉点上关键的中间代谢物而相 互影响和相互转化。这些共同的中间代谢物使各代谢途径得以沟通,形成经济有效、运转 良好的代谢网络。其中三个最关键的中间代谢物是6磷酸葡萄糖、 丙酮酸和乙酰辅酶A。 二、代谢的单向性和多酶系统 分解代谢和合成代谢可以是同一反应的逆转,但是事实上并不完全如此,整个代谢过 程都是单向的。例如,从葡萄糖降解成丙酮酸的酶共有十种,其中有三个酶参与的反应在 合成代谢过程中都是由别的酶或别的代谢途径所替代。也就是说在某些关键部位的正逆反 应往往是相对独立的单向反应,分别由不同的酶作化。这样可以从分解和合成两个方向分 别形成更有效的调节,在需要某一途径开放时另一途径就关闭,这种双重调节是代谢的普 遍现象。 细胞中的酶常常为了催化一系列连锁反应而联系成多酶系统,根据结构的复杂程度多 酶系统可分三种类型。第一种类型酶分子呈溶解状态,底物和产物一般均为小分子。扩散 速度快,易从一个酶分子扩散到另一个酶分子。第二种类型酶分子结构较紧密,各个酶分 子中间靠物理性质吸附,在功能上组成一个多酶系统,如脂肪酸合成酶系和丙酮酸脱氢酶 系,分别由7种和3种酶组合在一起,形城不易解离的复合体,一旦分开后就失去活性,底 物分子并不离开酶系,不扩散出去,可连续作用,提高了反应速率。第三种类型酶连接在 323
323 代谢物,如己糖、戊糖、生糖氨基酸和甘油降解为丙酮酸然后生成乙酰辅酶A,同样脂肪 酸和生酮氨基酸也降解为乙酰辅酶A和其他几个末端产物;最后阶段是乙酰辅酶A和其他产 物氧化成水和CO2。 合成代谢则经历反向的三个阶段,首先是以分解代谢第三阶段中生成的(或从环境中 摄入的)小分子作为起始原料合成简单有机物。不同的生物所利用的起始原料很不相同, 基本上分为二类,一类以二氧化碳为同化起始物合成各种有机物质(称为自养生物),不 需要由外界提供有机的碳化合物,如高等绿色植物、蓝绿藻、光合细菌、硝酸和亚硝酸细 菌等。另一类只能利用有机碳化物为起始原料(称为异养生物),如所有较高等的动物、 无光合作用的植物细胞、大多数微生物等。第二阶段是简单的有机物进一步合成构造大分 子所需要的单体。如α-酮酸与氨基供体反应生成α-氨基酸,磷酸丙糖生成磷酸己糖和磷 酸戊糖。丙酮酸经糖原异生作用生成的单酰乙酰辅酶A可缩合生成脂肪酸,磷酸二羟丙酮 可还原为甘油。第三阶段是由单体合成大分子化合物,如α-氨基酸结合成多肽;磷酸己糖 合成多糖;磷酸戊糖先形成核苷酸,然后合成核酸;甘油和脂肪酸合成脂肪。 分解代谢从各种不同的多糖、脂肪、蛋白质开始逐步形成共同的中间代谢物到第三阶 段则集中到一个共同的代谢途径三羧酸循环中。当然,在复杂的代谢中第二阶段的氨基酸、 甘油、单糖等单体并不是都要经过三羧酸循环,也可以直接作为合成大分子的中间物。合 成代谢则与之相反,一开始是共同的少数起始原料,然后逐步分化,经过第二阶段到第一 阶段产生了许多分枝途径,合成了许多不同类型的生物大分子。生物大分子具有高度的特 异性,正是它们决定了生物物种和个体之间的差异。生物机体将各类物质分别纳入各自的 共同代谢途径,以少数种类的反应,如氧化还原、基团转移、水解脱水、裂解合成、异构 反应等,转化成种类繁多的分子。不同的代谢途径可通过交叉点上关键的中间代谢物而相 互影响和相互转化。这些共同的中间代谢物使各代谢途径得以沟通,形成经济有效、运转 良好的代谢网络。其中三个最关键的中间代谢物是6-磷酸葡萄糖、丙酮酸和乙酰辅酶A。 二、代谢的单向性和多酶系统 分解代谢和合成代谢可以是同一反应的逆转,但是事实上并不完全如此,整个代谢过 程都是单向的。例如,从葡萄糖降解成丙酮酸的酶共有十种,其中有三个酶参与的反应在 合成代谢过程中都是由别的酶或别的代谢途径所替代。也就是说在某些关键部位的正逆反 应往往是相对独立的单向反应,分别由不同的酶催化。这样可以从分解和合成两个方向分 别形成更有效的调节,在需要某一途径开放时另一途径就关闭,这种双重调节是代谢的普 遍现象。 细胞中的酶常常为了催化一系列连锁反应而联系成多酶系统,根据结构的复杂程度多 酶系统可分三种类型。第一种类型酶分子呈溶解状态,底物和产物一般均为小分子。扩散 速度快,易从一个酶分子扩散到另一个酶分子。第二种类型酶分子结构较紧密,各个酶分 子中间靠物理性质吸咐,在功能上组成一个多酶系统,如脂肪酸合成酶系和丙酮酸脱氢酶 系,分别由7种和3种酶组合在一起,形成不易解离的复合体,一旦分开后就失去活性,底 物分子并不离开酶系,不扩散出去,可连续作用,提高了反应速率。第三种类型酶连接在

草上或核蛋白体上,如氧化电子传递链旅系,电子从底物传到氧,这些髓都在线拉体内榄 上。光合电子传递链酶系电子从HO传递到NADP,这些酶都在叶绿体的内囊体膜上。它 们就是膜结构的一部分。 另外,有一些酶能可逆地与膜结合,称之为双关酶(ambiguous enzyme)。它们有溶 解态和膜结合态二种形式。通过与膜的结合与解离迅速调节其活性,应答迅速、灵敏。如 糖酵解中的己糖激酶、氨基酸代谢中的谷氨酸脱氢酶以及一些参与蛋白质共价修饰的激酶 和磷酸(酯)酶等,是细跑代谢调节的重要方式之一。 三、代谢与能量 在分解代谢和合成代谢过程中都包括有能量的释放和利用。生物体内涉及到能量转换 的一切变化都遵循热力学的规律。一种需能反应与另一种放能反应联系在一起,使两个反 应都能进行,这就是活细胞中经常出现的偶联作用(coupling)。 有机体从环境中获得能量的方式各有不同,有的利用太阳的辐射能,有的利用氧化还 原反应中释放出来的化学能,不管哪种形式,细胞都能将它转变成高能分子ATP。ATP的 末端磷酸根的转换率很高,处于迅速代谢之中,在它水解脱去一个磷酸根时,大约释放出 2.9X10J/mol的能量。在完整细胞恒态情况下,ATP、ADP、AMP的浓度是相对稳定的, 可以用能荷的概念来表示细胞中腺苷酸库的高能磷酸化状态。其定义如下: 能荷= [ATP]+0.5[ADP] ATPI+ADPI+[AMPI 从理论上讲,能荷数值可以从零到1.0。但根据测定,大多数细胞中ATP的相对浓度较高 能荷值在0.8到0.9S之间。当细胞消耗较多的能量时,ATP的浓度下降,ADP浓度上升。能 荷稍有下降,立即促进糖酵解、三羧酸循环、氧化磷酸化和光合磷酸化过程,生成ATP。 反之,当ATP积累时,上述过程就被抑制,以降低ATP的生成速度。因为ATP的合成都是 由别构酶控制,ATP、ADP和AMP又往往是这些别构酶的调节物。所以胞内ATP水平较为 恒定,并不发生大幅度的变化。 放能和需能的反应相偶联,以ATP为中心 一方面有利于化学能的保存和利用,一方 面有利于代谢过程的顺利进行。生物合成往往是一种吸能反应或者是一种还原过程,它通 过与能量载体ATP的水解相偶联或者由还原型辅酶NADPH提供还原力(即氢和电子供体)。 分解代谢则是放能反应或者是一种氧化过程,生物体内除了ATP-ADP-AMP系统外,其他 的核苷酸也参与能量的 运转过程。这些核苷酸 与ATP之间可以互相转 换,共同参与生物大分 子的合成过程 A TP U TP A TP GIP AIP CIP 生物体内的高能化 多糖蛋白质 RNA DNA 合物除了核苷三磷酸 外,还有1,子二磷酸甘油酸中的酰基磷酸键。另外,如烯醇酯、硫酯和磷酸胍化合物,如
324 膜上或核蛋白体上,如氧化电子传递链酶系,电子从底物传到氧,这些酶都在线粒体内膜 上。光合电子传递链酶系电子从H2O传递到NADP+,这些酶都在叶绿体的内囊体膜上。它 们就是膜结构的一部分。 另外,有一些酶能可逆地与膜结合,称之为双关酶(ambiguous enzyme)。它们有溶 解态和膜结合态二种形式。通过与膜的结合与解离迅速调节其活性,应答迅速、灵敏。如 糖酵解中的己糖激酶、氨基酸代谢中的谷氨酸脱氢酶以及一些参与蛋白质共价修饰的激酶 和磷酸(酯)酶等,是细胞代谢调节的重要方式之一。 三、代谢与能量 在分解代谢和合成代谢过程中都包括有能量的释放和利用。生物体内涉及到能量转换 的一切变化都遵循热力学的规律。一种需能反应与另一种放能反应联系在一起,使两个反 应都能进行,这就是活细胞中经常出现的偶联作用(coupling)。 有机体从环境中获得能量的方式各有不同,有的利用太阳的辐射能,有的利用氧化还 原反应中释放出来的化学能,不管哪种形式,细胞都能将它转变成高能分子ATP。ATP的 末端磷酸根的转换率很高,处于迅速代谢之中,在它水解脱去一个磷酸根时,大约释放出 2.9×104 J/ mol的能量。在完整细胞恒态情况下,ATP、ADP、AMP的浓度是相对稳定的, 可以用能荷的概念来表示细胞中腺苷酸库的高能磷酸化状态。其定义如下: 能荷 [ATP] [ATP] [ADP] [AMP] [ADP] = +0.5 + + 从理论上讲,能荷数值可以从零到1.0。但根据测定,大多数细胞中ATP的相对浓度较高, 能荷值在0.8到0.95之间。当细胞消耗较多的能量时,ATP的浓度下降,ADP浓度上升。能 荷稍有下降,立即促进糖酵解、三羧酸循环、氧化磷酸化和光合磷酸化过程,生成ATP。 反之,当ATP积累时,上述过程就被抑制,以降低ATP的生成速度。因为ATP的合成都是 由别构酶控制,ATP、ADP和AMP又往往是这些别构酶的调节物。所以胞内ATP水平较为 恒定,并不发生大幅度的变化。 放能和需能的反应相偶联,以ATP为中心,一方面有利于化学能的保存和利用,一方 面有利于代谢过程的顺利进行。生物合成往往是一种吸能反应或者是一种还原过程,它通 过与能量载体ATP的水解相偶联或者由还原型辅酶NADPH提供还原力(即氢和电子供体)。 分解代谢则是放能反应或者是一种氧化过程,生物体内除了ATP-ADP-AMP系统外,其他 的核苷酸也参与能量的 运转过程。这些核苷酸 与ATP之间可以互相转 换,共同参与生物大分 子的合成过程。 生物体内的高能化 合物除了核苷三磷酸 外,还有1,3-二磷酸甘油酸中的酰基磷酸键。另外,如烯醇酯、硫酯和磷酸胍化合物,如
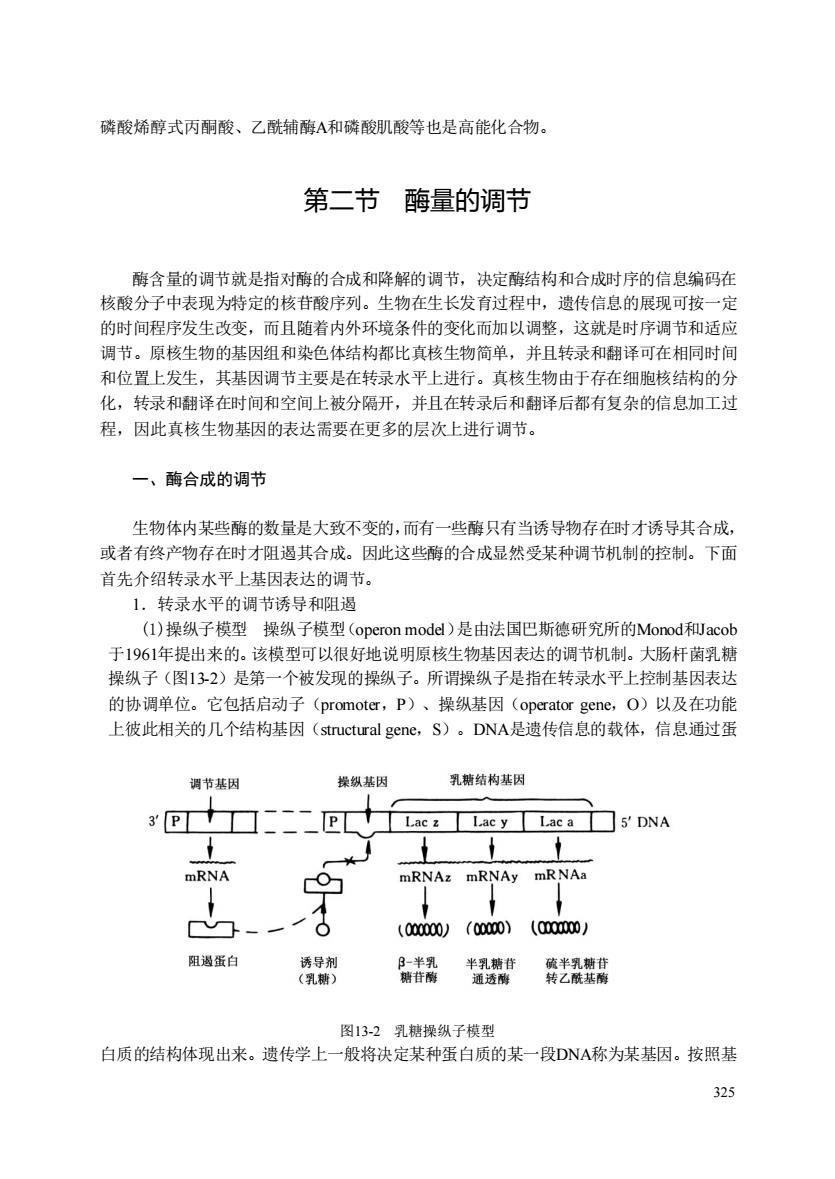
磷酸烯醇式丙酮酸、乙酰辅酶A和磷酸肌酸等也是高能化合物。 第二节酶量的调节 酶含量的调节就是指对酶的合成和降解的调节,决定酶结构和合成时序的信息编码在 核酸分子中表现为特定的核苷酸序列。生物在生长发育过程中,遗传信息的展现可按一定 的时间程序发生改变,而且随着内外环境条件的变化而加以调整,这就是时序调节和适应 调节。原核生物的基因组和染色体结构都比真核生物简单,并且转录和翻译可在相同时间 和位置上发生,其基因调节主要是在转录水平上进行。真核生物由于存在细胞核结构的分 化,转录和翻译在时间和空间上被分隔开,并且在转录后和翻译后都有复杂的信息加工过 程,因此真核生物基因的表达需要在更多的层次上进行调节。 一、酶合成的调节 生物体内某些酶的数量是大致不变的,而有一些酶只有当诱导物存在时才诱导其合成。 或者有终产物存在时才阻遏其合成。因此这些酶的合成显然受某种调节机制的控制。下面 首先介绍转录水平上基因表达的调节。 1.转录水平的调节诱导和阻遏 (I)操纵子模型操纵子模型(operon model)是由法国巴斯德研究所的Monod和acob 于1961年提出来的。该模型可以很好地说明原核生物基因表达的调节机制。大肠杆菌乳糖 操纵子(图132)是第一个被发现的操纵子。所谓操纵子是指在转录水平上控制基因表达 的协调单位。它包括启动子(promoter,P)、操纵基因(operator gene,.O)以及在功能 上彼此相关的几个结构基因(structural gene,S)。DNA是遗传信息的载体,信息通过蛋 调节基因 操纵基因 乳结构基 3E十☐二二二EL十Lac:Lacy Lac▣5DNA mRNA 0 RNAZ ORNAy mRNAa -0 (00000)(0000)(0000)/ 阻蛋白 半老辣 图132乳糖操纵子模型 白质的结构体现出来。遗传学上一般将决定某种蛋白质的某一段DNA称为某基因。按照基 325
325 磷酸烯醇式丙酮酸、乙酰辅酶A和磷酸肌酸等也是高能化合物。 第二节 酶量的调节 酶含量的调节就是指对酶的合成和降解的调节,决定酶结构和合成时序的信息编码在 核酸分子中表现为特定的核苷酸序列。生物在生长发育过程中,遗传信息的展现可按一定 的时间程序发生改变,而且随着内外环境条件的变化而加以调整,这就是时序调节和适应 调节。原核生物的基因组和染色体结构都比真核生物简单,并且转录和翻译可在相同时间 和位置上发生,其基因调节主要是在转录水平上进行。真核生物由于存在细胞核结构的分 化,转录和翻译在时间和空间上被分隔开,并且在转录后和翻译后都有复杂的信息加工过 程,因此真核生物基因的表达需要在更多的层次上进行调节。 一、酶合成的调节 生物体内某些酶的数量是大致不变的,而有一些酶只有当诱导物存在时才诱导其合成, 或者有终产物存在时才阻遏其合成。因此这些酶的合成显然受某种调节机制的控制。下面 首先介绍转录水平上基因表达的调节。 1.转录水平的调节诱导和阻遏 (1)操纵子模型 操纵子模型(operon model)是由法国巴斯德研究所的Monod和Jacob 于1961年提出来的。该模型可以很好地说明原核生物基因表达的调节机制。大肠杆菌乳糖 操纵子(图13-2)是第一个被发现的操纵子。所谓操纵子是指在转录水平上控制基因表达 的协调单位。它包括启动子(promoter,P)、操纵基因(operator gene,O)以及在功能 上彼此相关的几个结构基因(structural gene,S)。DNA是遗传信息的载体,信息通过蛋 图13-2 乳糖操纵子模型 白质的结构体现出来。遗传学上一般将决定某种蛋白质的某一段DNA称为某基因。按照基