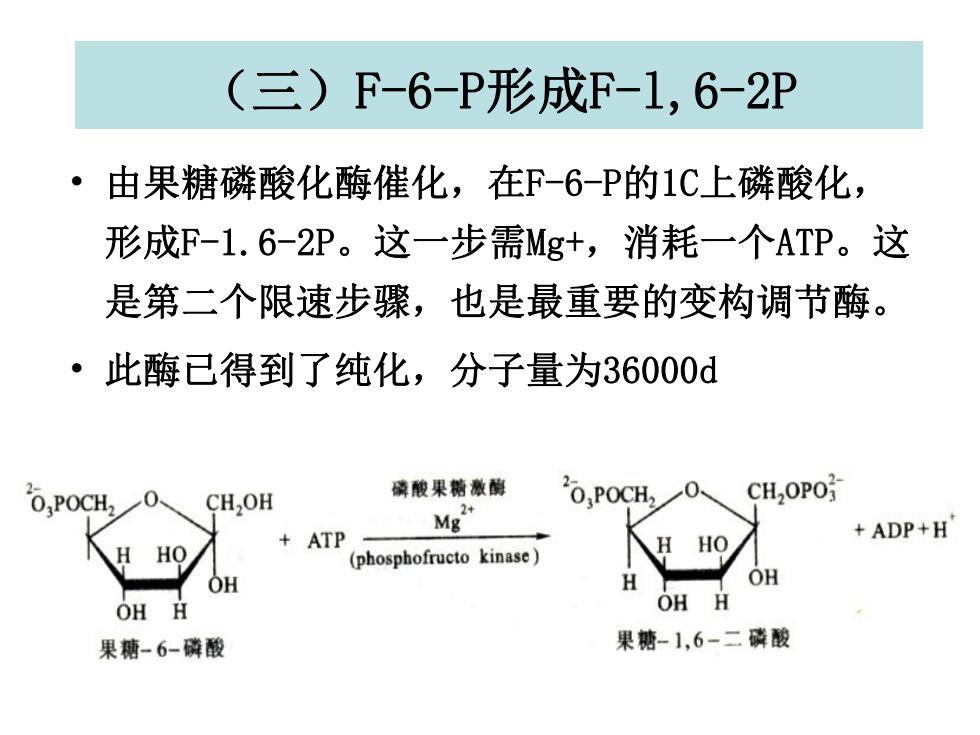
(三)F-6-P形成F-1,6-2P ·由果糖磷酸化酶催化,在F-6-P的1C上磷酸化, 形成F-1.6-2P。这一步需Mg+,消耗一个ATP。这 是第二个限速步骤,也是最重要的变构调节酶。 ·此酶已得到了纯化,分子量为36000d O,POCH, 磷酸果糖激脚 CH,OH 20,POCH CH2OPO +ATP ” +ADP+H HO (phosphofructo kinase) OH OH H OH H 果糖-6-磷酸 果糖-1,6-二磷酸
(三)F-6-P形成F-l,6-2P • 由果糖磷酸化酶催化,在F-6-P的1C上磷酸化, 形成F-1.6-2P。这一步需Mg+,消耗一个ATP。这 是第二个限速步骤,也是最重要的变构调节酶。 • 此酶已得到了纯化,分子量为36000d

(四)果糖-1,6-二磷酸转变为甘油醛-3-磷酸和 磷酸二羟丙酮 ·F-1,6-2P在醛缩酶的催化下,在C3-C4之间断裂形 成两个3C物。互为异构体,可互变。 醛缩酶有两种,醛缩酶I和醛缩酶Ⅱ,酶I存在于 动物细胞中,有4个亚基,Mt16万,3种同工酶。肌 肉型、肝脏型和大脑型。酶Ⅱ在微生物中,有2个 亚基,需Zn激活
(四)果糖-l,6-二磷酸转变为甘油醛-3-磷酸和 磷酸二羟丙酮 • F-1,6-2P在醛缩酶的催化下,在C3-C4之间断裂形 成两个3C物。互为异构体,可互变。 • 醛缩酶有两种,醛缩酶Ⅰ和醛缩酶Ⅱ,酶Ⅰ存在于 动物细胞中,有4个亚基,Mt16万,3种同工酶。肌 肉型、肝脏型和大脑型。酶Ⅱ在微生物中,有2个 亚基,需Zn激活

1CH2OPO 2C=0 1CH2OPO H.c-0 HO-C-H 醛缩酶 2C=0 + H-C-OH H7C-OH (aldolase) HOC-H 31 5 6CH,OPO3 HC-OH H 6CH2OPO 果糖-1,6-二磷酸 二羟丙酮磷酸 甘油醛-3-磷酸 (FBP) (DHAP) (GAP)

(五)磷酸二羟丙酮转变为甘油醛一3-磷酸 ·F-1,6-2P裂解后形成的两个3C糖,只有3-P-甘油 醛进入糖酵解途径,磷酸二羟丙酮在丙糖磷酸异 构酶催化下形成3-P-甘油醛。此酶有两个不同的 亚基,Mt56000。酶的催化是可逆的,但3-P-甘 油醛不断消耗,是反应总是向3-P-甘油醛方向进 行
(五)磷酸二羟丙酮转变为甘油醛-3-磷酸 • F-1,6-2P裂解后形成的两个3C糖,只有3-P-甘油 醛进入糖酵解途径,磷酸二羟丙酮在丙糖磷酸异 构酶催化下形成3-P-甘油醛。此酶有两个不同的 亚基,Mt56000。酶的催化是可逆的,但3-P-甘 油醛不断消耗,是反应总是向3-P-甘油醛方向进 行
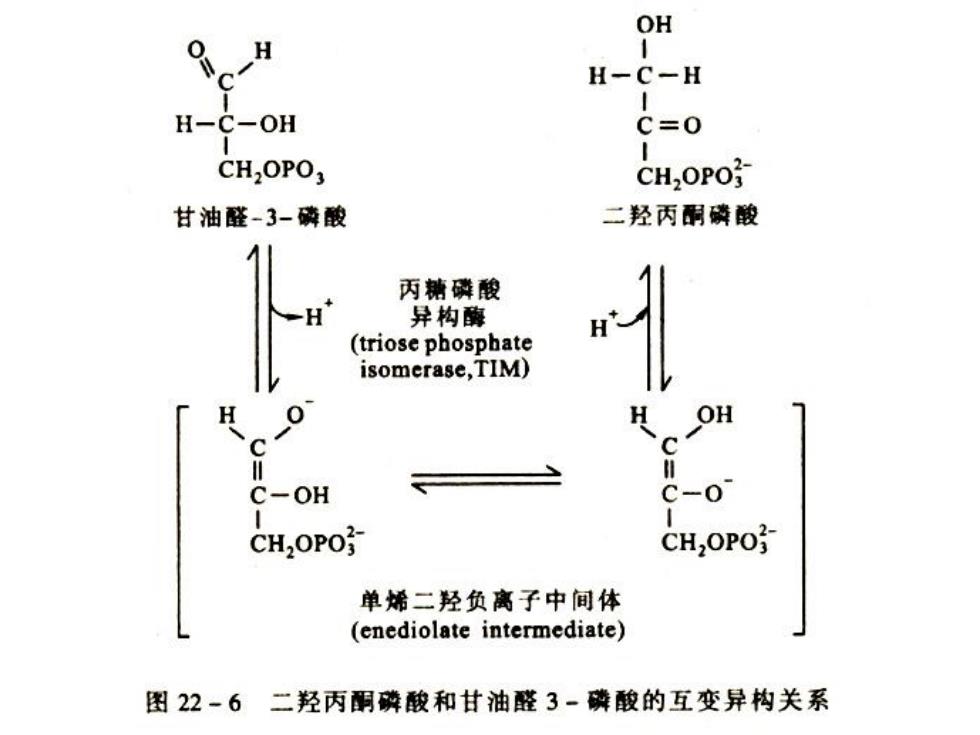
OH H-C-H H-C-OH C=0 CH,OPO; CH,OPO 甘油醛-3-磷酸 二羟丙酮磷酸 丙糖磷酸 异构酶 (triose phosphate isomerase,TIM) H OH C-OH CH2OPO CH,OPO 单烯二羟负离子中间体 (enediolate intermediate) 图22-6二羟丙酮磷酸和甘油醛3-磷酸的互变异构关系