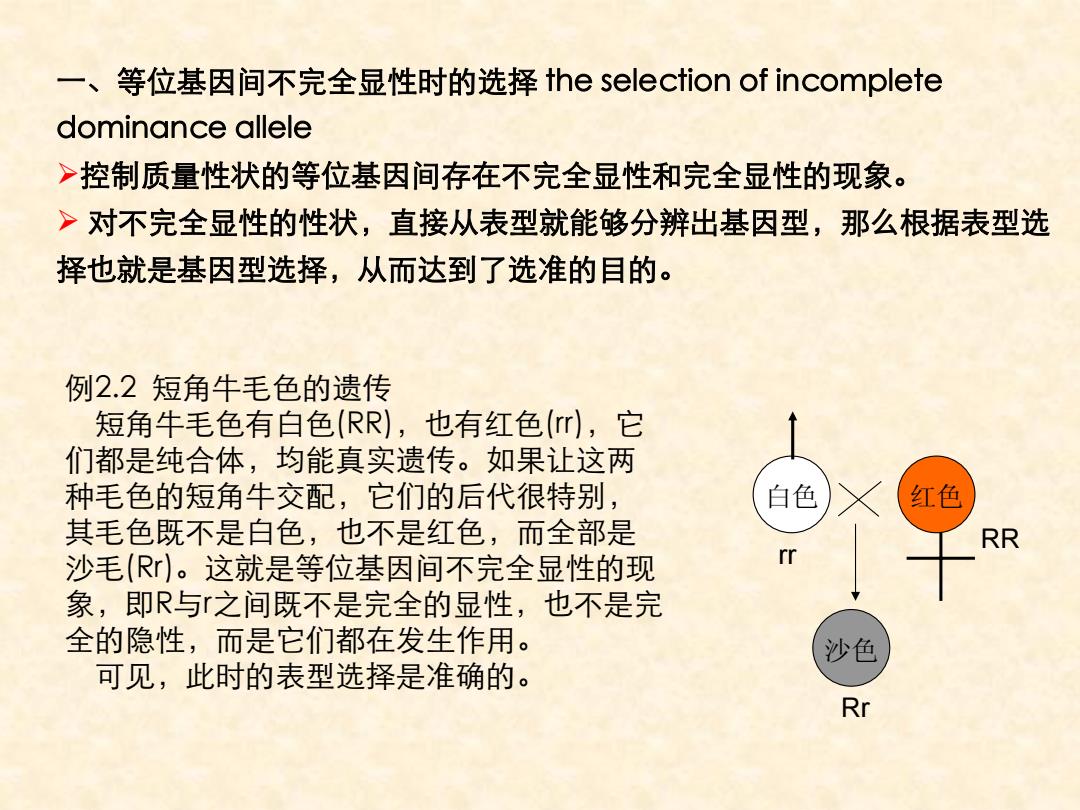
一、等位基因间不完全显性时的选择the selection of incomplete dominance allele >控制质量性状的等位基因间存在不完全显性和完全显性的现象。 >对不完全显性的性状,直接从表型就能够分辨出基因型,那么根据表型选 择也就是基因型选择,从而达到了选准的目的。 例2.2短角牛毛色的遗传 短角牛毛色有白色(RR),也有红色(r),它 们都是纯合体,均能真实遗传。如果让这两 种毛色的短角牛交配,它们的后代很特别, 白色 红色 其毛色既不是白色,也不是红色,而全部是 RR 沙毛(R)。这就是等位基因间不完全显性的现 rr 象,即R与之间既不是完全的显性,也不是完 全的隐性,而是它们都在发生作用。 沙色 可见,此时的表型选择是准确的。 Rr
一、等位基因间不完全显性时的选择 the selection of incomplete dominance allele ¾控制质量性状的等位基因间存在不完全显性和完全显性的现象。 ¾ 对不完全显性的性状,直接从表型就能够分辨出基因型,那么根据表型选 择也就是基因型选择,从而达到了选准的目的。 例2.2 短角牛毛色的遗传 短角牛毛色有白色(RR),也有红色(rr),它 们都是纯合体,均能真实遗传。如果让这两 种毛色的短角牛交配,它们的后代很特别, 其毛色既不是白色,也不是红色,而全部是 沙毛(Rr)。这就是等位基因间不完全显性的现 象,即R与r之间既不是完全的显性,也不是完 全的隐性,而是它们都在发生作用。 可见,此时的表型选择是准确的。 白色 红色 沙色 rr RR Rr

二、对隐性基因的选择 ·对隐性基因的选择实际上是对显性基因的淘汰过程 ·当显性基因的外显率是100%,且杂合子与显性纯合 子的表型相同时,则可以通过表型鉴别,一次性地 将显性基因全部淘汰 ·但一次性淘汰的做法会使部分“高产基因”随之丢失 ·明智的育种策略是,在保证生产性能不下降的前提 下,逐步完成对隐性基因的选择
二、对隐性基因的选择 •对隐性基因的选择实际上是对显性基因的淘汰过程 •当显性基因的外显率是100%,且杂合子与显性纯合 子的表型相同时,则可以通过表型鉴别,一次性地 将显性基因全部淘汰 •但一次性淘汰的做法会使部分 “高产基因 ”随之丢失 •明智的育种策略是,在保证生产性能不下降的前提 下,逐步完成对隐性基因的选择

确定显性纯合子及杂合子:首先借助等位基因间的显隐性关系,区分显 性个体与隐性个体,然后通过测交进一步识别显性纯合体与显性杂合 体,即判断其基因型。 当淘汰率为S时基因频率的基本计算: 设一对等位基因A、a,它们所组成的基因型为AA、Aa和aa。A对a完全显性; AA和Aa表型相同,为显性性状。若以相同的淘汰率淘汰公畜和母畜中的一部分 显性类型;交配方式的确定又与这一对性状无关,也就是对于这一对性状来说, 交配是随机的;以隐性基因频率的变化作为选择进展的标志,并设原始群体的基 因型频率为:D=0,片2p9,R=q2;淘汰率为S,(1-S即为留种率。于是得到 下表
确定显性纯合子及杂合子:首先借助等位基因间的显隐性关系,区分显 性个体与隐性个体,然后通过测交进一步识别显性纯合体与显性杂合 体,即判断其基因型。 当淘汰率为S时基因频率的基本计算: 设一对等位基因A、a,它们所组成的基因型为AA、Aa和aa。A对a完全显性; AA和Aa表型相同,为显性性状。若以相同的淘汰率淘汰公畜和母畜中的一部分 显性类型;交配方式的确定又与这一对性状无关,也就是对于这一对性状来说, 交配是随机的;以隐性基因频率的变化作为选择进展的标志,并设原始群体的基 因型频率为:D=p2,H=2pq,R=q2;淘汰率为s,(1- s)即为留种率。于是得到 下表

部分淘汰显性个体时选择前后基因型频率的变化 基因型 AA Aa aa 选择前频率 p 2p4 d 留种率 1-s 1-s 1 对下代贡献 p2(1-) 2pq(1-s) g 选择后频率 p2(1-s) 2pg(1-s) 92 1-s(1-g2) 1-s(1-g2) 1-s(1-92) 选择前基型频率X留种中率 选择后的基型频率= ∑选择前基型频率×留种摔 Σ(选择前基因型频率×留种率) =p2(1-S)+2pq(1-s)+q2 =(p2+2pq)(1-S)+g2 =(1-q2)(1-S)+q2 (.p2+2pq+q2=1) =1-s(1-q2) 注:上式中的分子可以理解为选择后各基因型的个体数,分母是淘汰率为S时 的选留总个体数
部分淘汰显性个体时选择前后基因型频率的变化 基因型 AA Aa aa 选择前频率 p 2 2pq q 2 留种率 1- s 1- s 1 对下代贡献 p 2(1- s ) 2pq(1- s ) q 2 选择后频率 p s s q 2 2 1 1 1 ( ) ( ) − − − 2 1 1 1 2 pq s s q ( ) ( ) − − − q s q 2 2 1 1 − − ( ) Σ(选择前基因型频率×留种率) =p2(1-s)+2pq(1-s)+q2 =(p2+2pq)(1-s)+q2 =(1-q2)(1-s)+q2 (∵p2+2pq+q2=1) =1-s(1-q2) 注:上式中的分子可以理解为选择后各基因型的个体数,分母是淘汰率为 s 时 的选留总个体数。 选择后的基因型频率 选择前基因型频率×留种率 选择前基因型频率×留种率 = ∑( )

选择后实行随机交配。设选择后的基因型频率分别为D←H”、和R‘,则下一代 的基因频率q1为: q1=1/2H+R’ 12pq(1-s) 92 21-5(1-92)1-s(1-92) =pg1-s)+g2-1-9)91-s)+92 1-5(1-q2) 1-s(1-92) =9-s(9-92) 1-s1-q2) 9=g-sg-q2) (2.2) 1-s(1-q2) 利用这个公式,根据对显性类型的淘汰率,就可以计算下一代的隐性基因频率。 9-q+g=1 当1时(淘汰全部显性个体),则: 1-1+q2 当=0时(没有淘汰显性个体),则:q1=q 这就是说当全部淘汰显性个体时,只要没有突变,下一代隐性基因的频率就可达 到1。可见选择隐性基因是相对容易的。但是,不完全淘汰显性个体时,下一代中 显性基因A还有一定的比例
选择后实行随机交配。设选择后的基因型频率分别为 D ‘ 、 H ’、和R‘,则下一代 的基因频率 q1为: q1=1/2H‘+R’ 利用这个公式,根据对显性类型的淘汰率,就可以计算下一代的隐性基因频率。 当 s=1时(淘汰全部显性个体),则: 当 s=0时(没有淘汰显性个体),则: q1= q 这就是说当全部淘汰显性个体时,只要没有突变,下一代隐性基因a的频率就可达 到1。可见选择隐性基因是相对容易的。但是,不完全淘汰显性个体时,下一代中 显性基因A还有一定的比例。 = ⋅ − − − + − − = − + − − = − −+ − − 1 2 2 1 11 11 1 1 1 1 1 1 1 2 2 2 2 2 2 2 pq s s q q s q pq s q s q qq s q s q ( ) () () ( ) ( ) ( )( ) ( ) = − − − − ∴ = − − − − q sq q s q q q sq q s q ( ) ( ) ( ) ( ) (.) 2 2 1 2 2 1 1 1 1 2 2 q qqq q 1 2 2 1 1 = 1 − + − + =