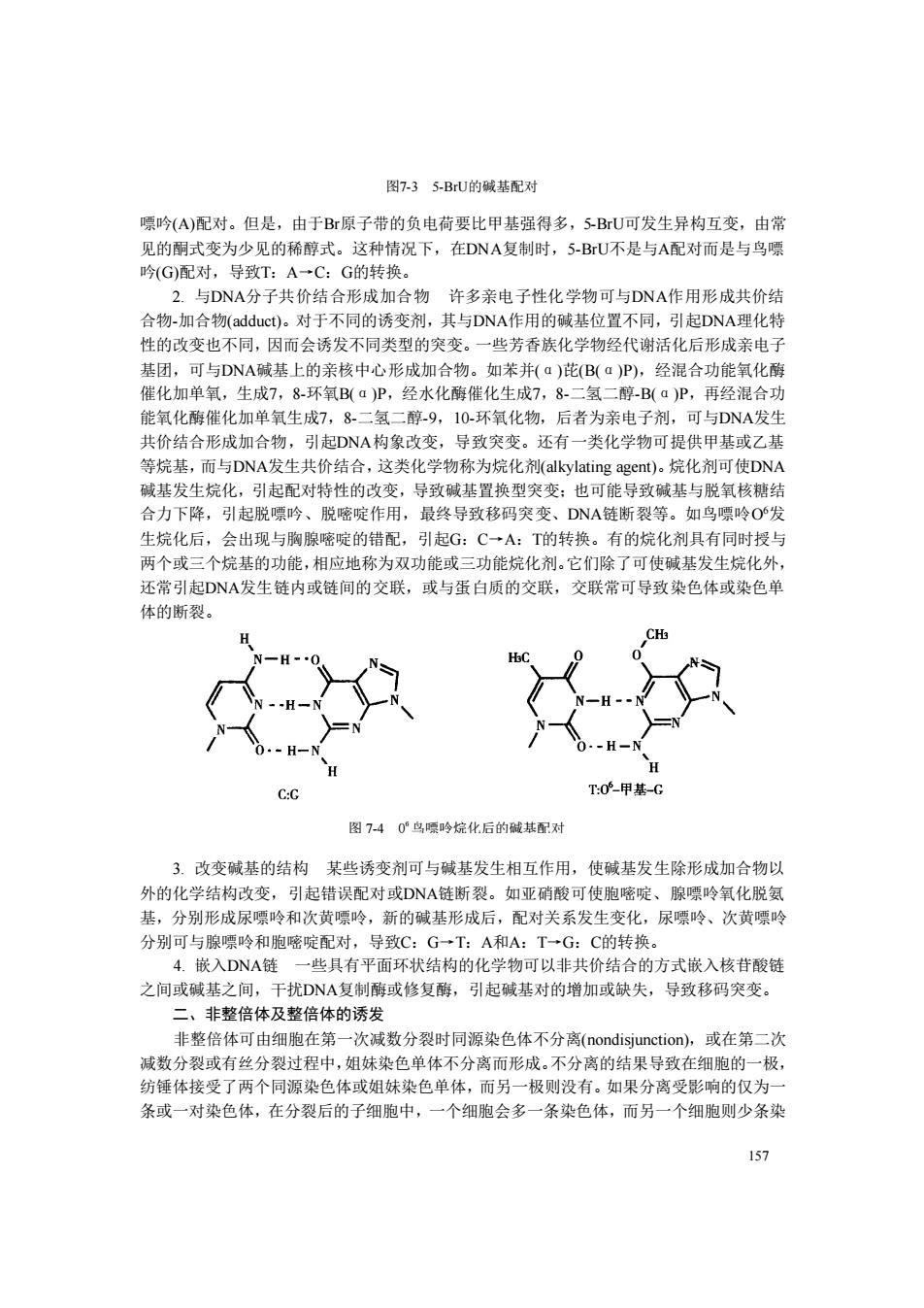
图7-35-BU的碱基配对 嘌吟(A)配对。但是,由于Br原子带的负电荷要比甲基强得多,5BU可发生异构互变,由常 见的租式变为少见的稀醇式。这种情况下,在DNA复制时,5-BU不是与A配对而是与鸟圆 吟(G)配对,导致T:A一C:G的转换 DNA分子共价结合形成加合物 许多亲电子性化学物可与DNA作用形成共价结 合物-加合物(adduct)。对于不同的诱变剂,其与DNA作用的碱基位置不同,引起DNA理化特 性的改变也不同,因而会诱发不同类型的突变。一些芳香族化学物经代谢活化后形成亲电子 基团,可与DNA碱基上的亲核中心形成加合物。如苯并(a)花Ba)P).经混合功能氧化 催化加单氧,生成7,8-环氧B叫aP,经水化酶催化生成78-二氢二醇-B(aP,再经混合功 能氧化酶催化加单氧生成7,8 一一破0 0环氧化物,后者为亲电子剂 可与DNA发生 共价结合形成加合物,引起DNA构象改变,导致突变。还有一类化学物可提供甲基或乙基 等烷基,而与DNA发生共价结合,这类化学物称为烷化剂(alkylating agent).烷化剂可使DNA 碱基发生烷化,引起配对特性的改变,导致碱基置换型突变:也可能导致碱基与脱氧核糖结 合力下降,引起脱嘌吟、脱嘧啶作用,最终导致移码突变、DNA链断裂等。如鸟嘌吟O发 生烷化后,会出现与胸腺嘧啶的错配,引起G:C一A:T的转换。有的烷化剂具有同时授与 两个或三个烷基的功能,相应地称为双功能或三功能烷化剂。它们除了可使碱基 生烷化列 还常引起D小A发生链内或链间的交联,或与蛋白质的交联,交联常可导致染色体或染色单 体的新裂。 0甲某C 图740心鸟暖吟烷化后的碱基配对 3.改变碱基的结构某些诱变剂可与碱基发生相互作用,使碱基发生除形成加合物以 外的化学结构改变,引起错误配对或DNA链断裂。如亚硝酸可使胞嘧啶、腺嘌呤氧化脱氨 基, 分别形成尿噤吟和次黄吟,新的碱基形成后,配对关系发生变化 尿嘌呤、次黄嘌吟 分别可与腺嘌吟和胞嘧啶配对,导致C:G一T:A和A:T一G:C的转换。 4.嵌入DNA链一些具有平面环状结构的化学物可以非共价结合的方式嵌入核苷酸链 之间或碱基之间,干扰DNA复制酶或修复酶,引起碱基对的增加或缺失,导致移码突变。 二、非整倍体及整倍体的诱发 非整倍体可由细胞在第 一次减数分裂时同源染色体不分离(nondisjunction),或在第二次 减数分裂或有丝分裂过程中,姐妹染色单体不分离而形成。不分离的结果导致在细胞的一极 纺锤体接受了两个同源染色体或姐妹染色单体,而另一极则没有。如果分离受影响的仅为 条或一对染色体,在分裂后的子细胞中,一个细胞会多一条染色体,而另一个细胞则少条染 157
157 图7-3 5-BrU的碱基配对 嘌吟(A)配对。但是,由于Br原子带的负电荷要比甲基强得多,5-BrU可发生异构互变,由常 见的酮式变为少见的稀醇式。这种情况下,在DNA复制时,5-BrU不是与A配对而是与鸟嘌 吟(G)配对,导致T:A→C:G的转换。 2. 与DNA分子共价结合形成加合物 许多亲电子性化学物可与DNA作用形成共价结 合物-加合物(adduct)。对于不同的诱变剂,其与DNA作用的碱基位置不同,引起DNA理化特 性的改变也不同,因而会诱发不同类型的突变。一些芳香族化学物经代谢活化后形成亲电子 基团,可与DNA碱基上的亲核中心形成加合物。如苯并(α)芘(B(α)P),经混合功能氧化酶 催化加单氧,生成7,8-环氧B(α)P,经水化酶催化生成7,8-二氢二醇-B(α)P,再经混合功 能氧化酶催化加单氧生成7,8-二氢二醇-9,10-环氧化物,后者为亲电子剂,可与DNA发生 共价结合形成加合物,引起DNA构象改变,导致突变。还有一类化学物可提供甲基或乙基 等烷基,而与DNA发生共价结合,这类化学物称为烷化剂(alkylating agent)。烷化剂可使DNA 碱基发生烷化,引起配对特性的改变,导致碱基置换型突变;也可能导致碱基与脱氧核糖结 合力下降,引起脱嘌吟、脱嘧啶作用,最终导致移码突变、DNA链断裂等。如鸟嘌呤O6发 生烷化后,会出现与胸腺嘧啶的错配,引起G:C→A:T的转换。有的烷化剂具有同时授与 两个或三个烷基的功能,相应地称为双功能或三功能烷化剂。它们除了可使碱基发生烷化外, 还常引起DNA发生链内或链间的交联,或与蛋白质的交联,交联常可导致染色体或染色单 体的断裂。 3. 改变碱基的结构 某些诱变剂可与碱基发生相互作用,使碱基发生除形成加合物以 外的化学结构改变,引起错误配对或DNA链断裂。如亚硝酸可使胞嘧啶、腺嘌呤氧化脱氨 基,分别形成尿嘌呤和次黄嘌呤,新的碱基形成后,配对关系发生变化,尿嘌呤、次黄嘌呤 分别可与腺嘌呤和胞嘧啶配对,导致C:G→T:A和A:T→G:C的转换。 4. 嵌入DNA链 一些具有平面环状结构的化学物可以非共价结合的方式嵌入核苷酸链 之间或碱基之间,干扰DNA复制酶或修复酶,引起碱基对的增加或缺失,导致移码突变。 二、非整倍体及整倍体的诱发 非整倍体可由细胞在第一次减数分裂时同源染色体不分离(nondisjunction),或在第二次 减数分裂或有丝分裂过程中,姐妹染色单体不分离而形成。不分离的结果导致在细胞的一极, 纺锤体接受了两个同源染色体或姐妹染色单体,而另一极则没有。如果分离受影响的仅为一 条或一对染色体,在分裂后的子细胞中,一个细胞会多一条染色体,而另一个细胞则少条染 图 7-4 0 6 鸟嘌呤烷化后的碱基配对

色体,分别形成三体和缺体。非整倍体剂(aneugen)有多种机制导致细胞分裂异常,诱发非整 倍体。其作用的靶可以为:①微管的合成和组装、纺锤体形成:②中心粒和极体的合成、分 裂及其功能:②着丝粒蛋白的组装及其功能和着丝粒DN 性细胞减数分裂与有丝分裂的 机理不同, 其非整倍体形成的机制也有所不同,所以,利用体细胞非整倍体试验不能检测对 减数分裂特异的非整倍体剂。 多倍体涉及整个染色体组。在有丝分裂过程中,若染色体己正常复制,但由于纺锤体受 损,垫色单体不能分离到子细胞中,这时染色体数目就会加倍,形成四倍体。成数分裂的异 常也可使配子形成二倍体,若二倍体的配子受精,可形成多倍体的受精卵。 一个卵子被多个 精子受精,也可形成多倍体 三、DNA损伤修复与突变 L.生物体对DNA损伤的修复及耐受 环境因素可引起各种类型的DNA损伤,并不是所有损伤都会表现为突变。细跑对于DNA 损伤有修复及耐受机制。细胞内G/S交界处的关卡点(checkPoint))负责检查染色体DA是否 有损伤,如果DNA有损伤 则把细胞阻止子 更求细先进 修复 后 能复制 免遗传信息出错。DNA受损后,细胞利用其修复系统对损伤进行修复,如果DNA损伤能被 正确无误地修复,那么,对生物体而言,这种损伤不会产生什么后果,亦即不引起突变。只 有损伤不能被修复或在修复中出现了错误,一般需经过2次或多次细胞周期才有可能固定下 来,并传递到后代的细胞或个体,引起突变。所以,环境致突变作用的模式应为遗传机构损 伤损伤修复·突变 周定突变 DNA损伤修复机制可分为直接修复(direct repair)和切除修复(excision repair))。直接修复 使损伤DNA回复正常,包括光修复及O户.甲基鸟嘌吟修复等。光修复是针对紫外线引起的晒 啶二聚体的修复功能,其修复机制比较简单,且很特异。在可见光存在的条件下,经酶的作 用将二聚体打开,使相邻的腔碱基回同复原来的结构,一般为无误修复。生物进化程度战高 此种修复功能越弱。O心.甲基鸟吟修复,修复在O位上含有烷基的鸟嘌吟,靠O.甲基鸟漂 吟-.DNA甲基转移酶将O5.甲基鸟嘌吟的甲基转移 至该 的半胱氨 酸残基上 而恢复鸟嘌 正常的碱基配对特性,该酶在修复过程中被不可逆地失活。在大肠杆菌、酵母、呐齿类及人 类细胞都发现有O甲基鸟嘌吟-DNA甲基转移酶。该酶具有可诱导性,如在大肠杆菌每个 细胞含13~60个醇分子,诱导后可增加到每个细胞3000个酶分子。对于其他的烷化碱基也可 能存在类似的特异修复系统。 切除修复指除去损伤碱基 、含损伤的DNA片段或错配碱基的机制。与光修复及O甲基 鸟嘌吟修复不同,该修复机制适应于广泛的DNA损伤类型,是最主要的DNA损伤修复途径 一般为无误修复。依据其切除对象的不同可分为核苷酸切除修复、碱基切除修复及错配碱基 修复三种类型。 核苷酸切除修复(ucleotide excision repair)是所有生物体内最常见的修复机制。它可修复 几乎所有的DNA损伤类型,包括其他修复机制不能修复的加合物及DNA链间交联等。修复 时,先靠内切酶把DNA链从损伤两端切断:在解螺旋酶helicase)作用下,除去受损的寡核苷 酸:再在修复多聚酶的作用下,以对应的链为模板,以正确的碱基填补空隙:最后,在DNA 连接酶的作用下连接,恢复原来序列。核苷酸切除修复时,对于存在于活性转录链上的嘧喷 二聚体和某些加合物等损伤比存在于互补的非转录链或无活性序列上的损伤可优先去除。对
158 色体,分别形成三体和缺体。非整倍体剂(aneugen)有多种机制导致细胞分裂异常,诱发非整 倍体。其作用的靶可以为:①微管的合成和组装、纺锤体形成;②中心粒和极体的合成、分 裂及其功能;②着丝粒蛋白的组装及其功能和着丝粒DNA。性细胞减数分裂与有丝分裂的 机理不同,其非整倍体形成的机制也有所不同,所以,利用体细胞非整倍体试验不能检测对 减数分裂特异的非整倍体剂。 多倍体涉及整个染色体组。在有丝分裂过程中,若染色体己正常复制,但由于纺锤体受 损,染色单体不能分离到子细胞中,这时染色体数目就会加倍,形成四倍体。减数分裂的异 常也可使配子形成二倍体,若二倍体的配子受精,可形成多倍体的受精卵。一个卵子被多个 精子受精,也可形成多倍体。 三、DNA损伤修复与突变 1. 生物体对DNA损伤的修复及耐受 环境因素可引起各种类型的DNA损伤,并不是所有损伤都会表现为突变。细胞对于DNA 损伤有修复及耐受机制。细胞内Gl/S交界处的关卡点(checkPoint)负责检查染色体DNA是否 有损伤,如果DNA有损伤,则把细胞阻止于Gl期,要求细胞先进行修复,然后才能复制,以 免遗传信息出错。DNA受损后,细胞利用其修复系统对损伤进行修复,如果DNA损伤能被 正确无误地修复,那么,对生物体而言,这种损伤不会产生什么后果,亦即不引起突变。只 有损伤不能被修复或在修复中出现了错误,一般需经过2次或多次细胞周期才有可能固定下 来,并传递到后代的细胞或个体,引起突变。所以,环境致突变作用的模式应为遗传机构损 伤-损伤修复-突变固定-突变。 DNA损伤修复机制可分为直接修复(direct repair)和切除修复(excision repair)。直接修复 使损伤DNA回复正常,包括光修复及O6 -甲基鸟嘌呤修复等。光修复是针对紫外线引起的嘧 啶二聚体的修复功能,其修复机制比较简单,且很特异。在可见光存在的条件下,经酶的作 用将二聚体打开,使相邻的嘧啶碱基回复原来的结构,一般为无误修复。生物进化程度越高, 此种修复功能越弱。O6 -甲基鸟嘌呤修复,修复在O6位上含有烷基的鸟嘌呤,靠O6 -甲基鸟漂 吟-DNA-甲基转移酶将O6 -甲基鸟嘌呤的甲基转移至该酶的半胱氨酸残基上,而恢复鸟嘌呤 正常的碱基配对特性,该酶在修复过程中被不可逆地失活。在大肠杆菌、酵母、啮齿类及人 类细胞都发现有O6 -甲基鸟嘌呤-DNA-甲基转移酶。该酶具有可诱导性,如在大肠杆菌每个 细胞含13~60个酶分子,诱导后可增加到每个细胞3000个酶分子。对于其他的烷化碱基也可 能存在类似的特异修复系统。 切除修复指除去损伤碱基、含损伤的DNA片段或错配碱基的机制。与光修复及O6甲基 鸟嘌呤修复不同,该修复机制适应于广泛的DNA损伤类型,是最主要的DNA损伤修复途径, 一般为无误修复。依据其切除对象的不同可分为核苷酸切除修复、碱基切除修复及错配碱基 修复三种类型。 核苷酸切除修复(nucleotide excision repair)是所有生物体内最常见的修复机制。它可修复 几乎所有的DNA损伤类型,包括其他修复机制不能修复的加合物及DNA链间交联等。修复 时,先靠内切酶把DNA链从损伤两端切断;在解螺旋酶(helicase)作用下,除去受损的寡核苷 酸;再在修复多聚酶的作用下,以对应的链为模板,以正确的碱基填补空隙;最后,在DNA 连接酶的作用下连接,恢复原来序列。核苷酸切除修复时,对于存在于活性转录链上的嘧啶 二聚体和某些加合物等损伤比存在于互补的非转录链或无活性序列上的损伤可优先去除。对

这种在修复时优先性的进一步深入研究有助于揭示转录、修复和突变的关系及这种关系在致 突变危险性评价中的意义。 碱基切除修复(repair)通常修补的是单个被损伤的核苷酸。由DNA糖基化酶 识别结构有改变的受 碱基,并通过水解其与脱氧核糖连接的健将损伤的碱基切除,形成 嘌吟/脱嘧啶(AP)位点,然后,AP内切酶将DNA链切断,并去除原来与受损碱基连接的脱 氧核糖,再在DNA聚合酶、连接酶的作用下填补失去碱基的部位,完成修复过程。DNA糖 基化酶比核苷酸切除酶具有更大的特异性,但仍可切除紫外线引起的嘧啶二聚体,辐射、烷 化剂、过氧化物等引起的不太大的损伤及诸如胞嘧啶脱氨基变为尿嘧啶的自发损伤等。 种特殊的切除修复形式 通过该机制可去除不 正确的碱基配对,如G:T和A:C。错配碱 基对可由复制时发生错误作为重组中间体出现 也可由碱基化学修饰形成。如果错配碱基对维持到下一个复制周明,将按正常的碱基配对关 系配对,在两个新的DNA分子中,一个分子正常,另一个则会含有一对错误的碱基。细胞 一般可以检查到错配碱基的存在,并讲行修复。修复时针对那一条练上的碱基,关系到是否 达到无误修复。如果错配是在复制过程中产生, 一般是对新合成的链进行修复。对于老链利 新链的识别,可根 DNA链的甲基化水平 新合成链处于低甲基化状态 除修复错配碱 外,该修复机制还可修复在DNA复制时形成的小的缺失及插入。错配修复的缺失将导致遗 传的不稳定性,错配碱基修复与肿瘤发生的关系是近年来人们关注的热点,已发现错配修复 的有关基因突变与遗传性非息肉性大肠癌的发生有关。 DNA损伤修复机制具有饱和性,另外,对于某些损伤也不能有效修复。没有被修复的 损伤, 维持到下 复制周期, 有些会影响依赖DNA多聚酶的复制的精确性 引起 变的发 生:另一些损伤可阻断DNA的复制,危及细胞的生存,此时,细胞可通过其耐受机制重新 启动处于复制阻断状态模板的DNA合成。一种途径是通过复制后修复的机制,复制时新合 成的互补链相应部位出现空隙,随后以重组作用及链延长作用筑补。通过此机制,使DNA 合成得以通过损伤部位,避免了致死性的后果,但存在的DNA损伤并末移除,仍有待通过 其他的修复机制而修复。另外 在大肠杆菌 ,阻止DNA复制的损伤等可诱发SOS修复系统, 即诱导细胞产生特殊的DNA聚合酶,以不严格的碱基配对使复制通过损伤部位。通过SOS 修复,细胞得以存活,但在此过程中常导入错误的碱基,故常为易误修复。 2.DNA损伤修复与突变 突变的产生不仅与DNA受损的情况有关,DNA损伤修复也是决定突变发生与否的重要 因素。DNA损伤修复功能的缺失或修复能力降低都会使突变的发生明显增加。如在有切除 修复缺的人成纤维细胞 对于紫外线引起突变的敏感性大大高于正常的成纤维细胞 表7-2太阳光照射诱发M13mD2噬茵体基因突变在不同宿主中的表达 突变頓率(×10 照射时间(min) 末诱导sOS功能宿主黄 透导SOS功能宿主黄 13.8 9.6 650 180 13.2 1002 159
159 这种在修复时优先性的进一步深入研究有助于揭示转录、修复和突变的关系及这种关系在致 突变危险性评价中的意义。 碱基切除修复(base excision repair)通常修补的是单个被损伤的核苷酸。由DNA糖基化酶 识别结构有改变的受损碱基,并通过水解其与脱氧核糖连接的键将损伤的碱基切除,形成脱 嘌呤/脱嘧啶(AP)位点,然后,AP内切酶将DNA链切断,并去除原来与受损碱基连接的脱 氧核糖,再在DNA聚合酶、连接酶的作用下填补失去碱基的部位,完成修复过程。DNA糖 基化酶比核苷酸切除酶具有更大的特异性,但仍可切除紫外线引起的嘧啶二聚体,辐射、烷 化剂、过氧化物等引起的不太大的损伤及诸如胞嘧啶脱氨基变为尿嘧啶的自发损伤等。 错配碱基修复(mismatch base repair)是一种特殊的切除修复形式,通过该机制可去除不 正确的碱基配对,如G:T和A:C。错配碱基对可由复制时发生错误作为重组中间体出现, 也可由碱基化学修饰形成。如果错配碱基对维持到下一个复制周期,将按正常的碱基配对关 系配对,在两个新的DNA分子中,一个分子正常,另一个则会含有一对错误的碱基。细胞 一般可以检查到错配碱基的存在,并进行修复。修复时针对那一条链上的碱基,关系到是否 达到无误修复。如果错配是在复制过程中产生,一般是对新合成的链进行修复。对于老链和 新链的识别,可根据DNA链的甲基化水平,新合成链处于低甲基化状态。除修复错配碱基 外,该修复机制还可修复在DNA复制时形成的小的缺失及插入。错配修复的缺失将导致遗 传的不稳定性,错配碱基修复与肿瘤发生的关系是近年来人们关注的热点,已发现错配修复 的有关基因突变与遗传性非息肉性大肠癌的发生有关。 DNA损伤修复机制具有饱和性,另外,对于某些损伤也不能有效修复。没有被修复的 损伤,维持到下一复制周期,有些会影响依赖DNA多聚酶的复制的精确性,引起突变的发 生;另一些损伤可阻断DNA的复制,危及细胞的生存,此时,细胞可通过其耐受机制重新 启动处于复制阻断状态模板的DNA合成。一种途径是通过复制后修复的机制,复制时新合 成的互补链相应部位出现空隙,随后以重组作用及链延长作用填补。通过此机制,使DNA 合成得以通过损伤部位,避免了致死性的后果,但存在的DNA损伤并末移除,仍有待通过 其他的修复机制而修复。另外,在大肠杆菌,阻止DNA复制的损伤等可诱发SOS修复系统, 即诱导细胞产生特殊的DNA聚合酶,以不严格的碱基配对使复制通过损伤部位。通过SOS 修复,细胞得以存活,但在此过程中常导入错误的碱基,故常为易误修复。 2. DNA损伤修复与突变 突变的产生不仅与DNA受损的情况有关,DNA损伤修复也是决定突变发生与否的重要 因素。DNA损伤修复功能的缺失或修复能力降低都会使突变的发生明显增加。如在有切除 修复缺陷的人成纤维细胞,对于紫外线引起突变的敏感性大大高于正常的成纤维细胞。 表7-2 太阳光照射诱发M13mp2噬茵体基因突变在不同宿主中的表达 照射时间(min) 突变频率(×10-4 ) 末诱导SOS功能宿主菌 诱导SOS功能宿主菌 0 1.8 13.8 60 9.6 65.0 180 13.2 100.2