
结论: ●加热杀死的SⅢ型细菌可以使活的RⅡ型细 菌合成SⅢ型荚膜多糖而成为有毒细菌,这 种现象叫转化(transformation) ●10年后Avery等的体外转化实验,弄清转化 因子是DNA
结论: ⚫加热杀死的SⅢ型细菌可以使活的RⅡ 型细 菌合成SⅢ型荚膜多糖而成为有毒细菌,这 种现象叫转化(transformation)。 ⚫10年后Avery等的体外转化实验,弄清转化 因子是DNA
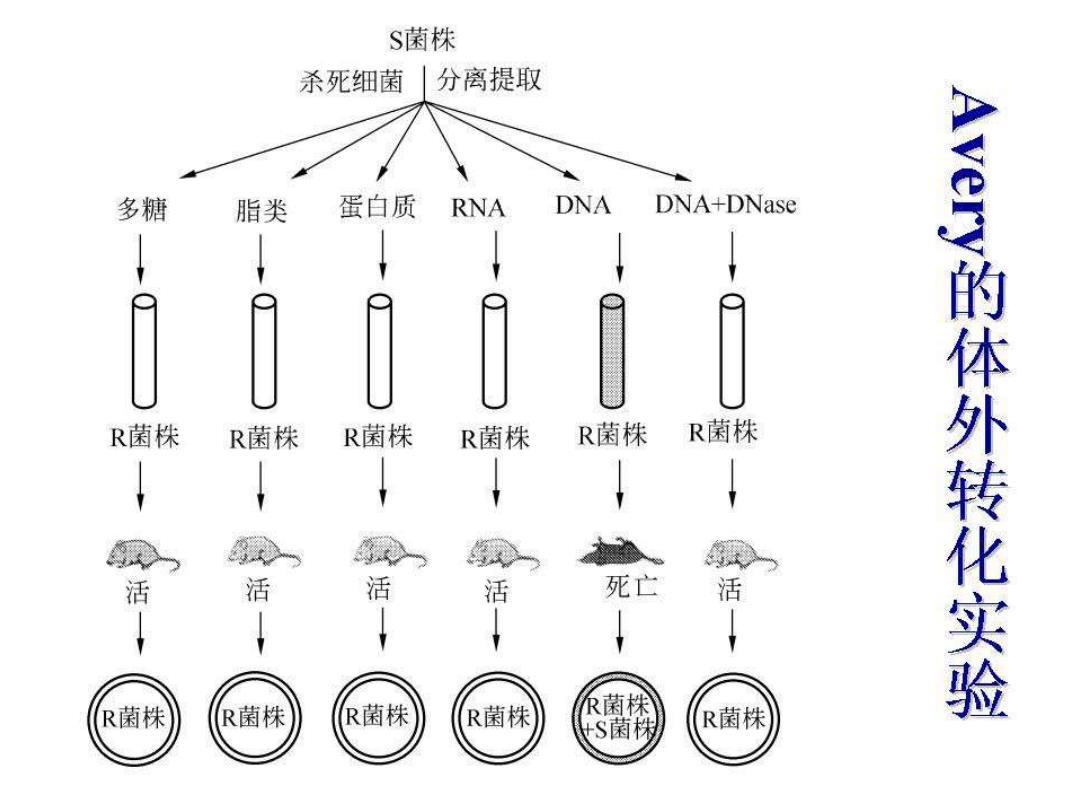
S菌株 杀死细菌分离提取 多糖 脂类 蛋白质 RNA DNA DNA+DNase R菌株 R菌株 R菌株 R菌株 R菌株 R菌株 活 活 活 活 死亡 活 e的体外转化实验 R菌株 R菌株 R菌株 R菌株 ?菌株 S菌 R菌株

二、噬菌体的侵染 噬菌体是微小的细菌病毒,其中T,噬菌体 侵染大肠杆菌。绝大多数是含有DNA的病 毒,只有少数含有RNA的病毒。 ●1952年赫尔歇(Hershey)和索斯 (Chose)进行噬菌体侵染的分析实验如 图:
二、噬菌体的侵染 ⚫噬菌体是微小的细菌病毒,其中T2噬菌体 侵染大肠杆菌。绝大多数是含有DNA的病 毒,只有少数含有RNA的病毒。 ⚫1952年赫尔歇(Hershey)和索斯 (Chose)进行噬菌体侵染的分析实验 如 图:

● 1952年,赫希和蔡斯把宿主细菌分别培养在含有35S和32P的培养基 1,宿主细菌在生长过程中就分别被3$和32P所标记。T2噬菌体由 壳蛋直和DNA组成,蛋质含有而不含有P,DNA含有而不 。赫希等人用T:噬菌还分脚長染被35和32P标记的细菌。,【 有 体在细菌的内增殖裂解后释 致出的子代噬菌体中1 3S标花细菌的噬園体蛋首液棕花,侵类32P标记细崮的噬菌体bN八 被标。接下来用被35$和32P记的噬菌体分别一 培养基 的细菌(培养1Qm后,经型烈薹荡使吸的娃菌体表面的城芬脱落 下 离父后脱落感汾悬浮在上清液中,菌连细胞沉迂底部 :d漾面躇禁 测 累雀威赛 镇我再 胞的同位素标记。 信 感 染的细菌测定军 远示,{ 素标记天多数同位32P标定信号出现美细胞内鹰1二2。 实验说明应体在得类细黄时进人细丙的主要分冕中 - DNA一附羞专到菌表面的是蛋名质。,司觅,在碰菌林的生结 DNA是起增殖产生字代的畅质。因此,再次证萌5NA是遗传物质
⚫ 1952年,赫希和蔡斯把宿主细菌分别培养在含有35S和32P的培养基 中,宿主细菌在生长过程中就分别被35S和32P所标记。T2噬菌体由 外壳蛋白和DNA组成,蛋白质含有S而不含有P, DNA含有P而不含 有S。赫希等人用T:噬菌体分别去侵染被35S和32P标记的细菌。噬 菌体在细菌细胞内增殖,细菌裂解后释放出的子代噬菌体中,侵染 35S标记细菌的噬菌体蛋白被标记,侵染32P标记细菌的噬菌体DNA 被标记。接下来用被35S和32P标记的噬菌体分别去侵染一般培养基 中的细菌,培养10min后,经剧烈震荡使吸附于菌体表面的成分脱落 下来,离心后,脱落成分悬浮在上清液中,菌体细胞沉于底部。测定 宿主细胞的同位素标记。对于用35S标记的噬菌体侵染的细菌,测定 结果显示,宿主细胞内很少有同位素标记,而大多数35S标记信号出 现在含有宿主细胞表面附着物的上清液里面;对于用32P标记的噬菌 体感染的细菌,测定结果显示,宿主细胞的表面附着物中很少有同位 素标记,而大多数同位素32P标记信号出现在宿主细胞内(图1—2)。 这一实验说明,噬菌体在侵染细菌时,进入细菌内的主要成分是 DNA,而附着于细菌表面的是蛋白质。可见,在噬菌体的生活史中, DNA是引起增殖产生子代的物质。因此,再次证明DNA是遗传物质
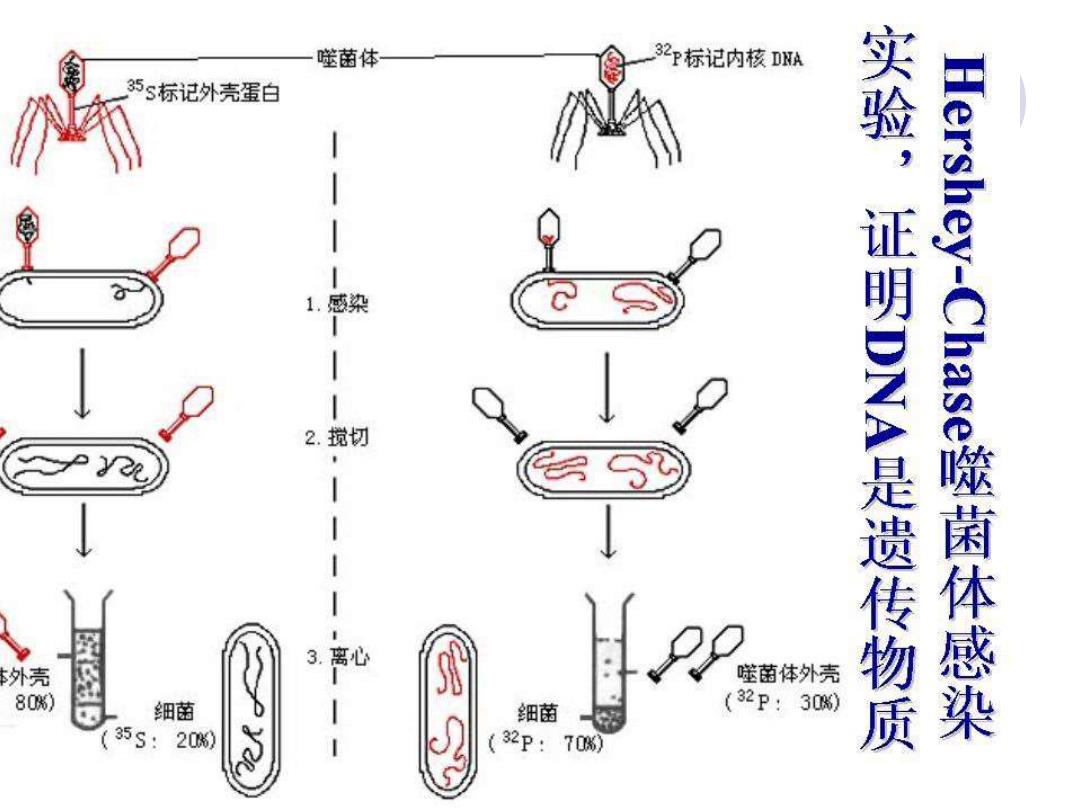
噬菌体 2P标记内核A 5s标记外壳蛋白 2 实验,证明02是遗传物质 隐菌体感染H 3 离心 外壳 噬菌体外壳 80%) 细菌 细菌 (32P:30瑞) 35S: 20%) (32p: 70%)