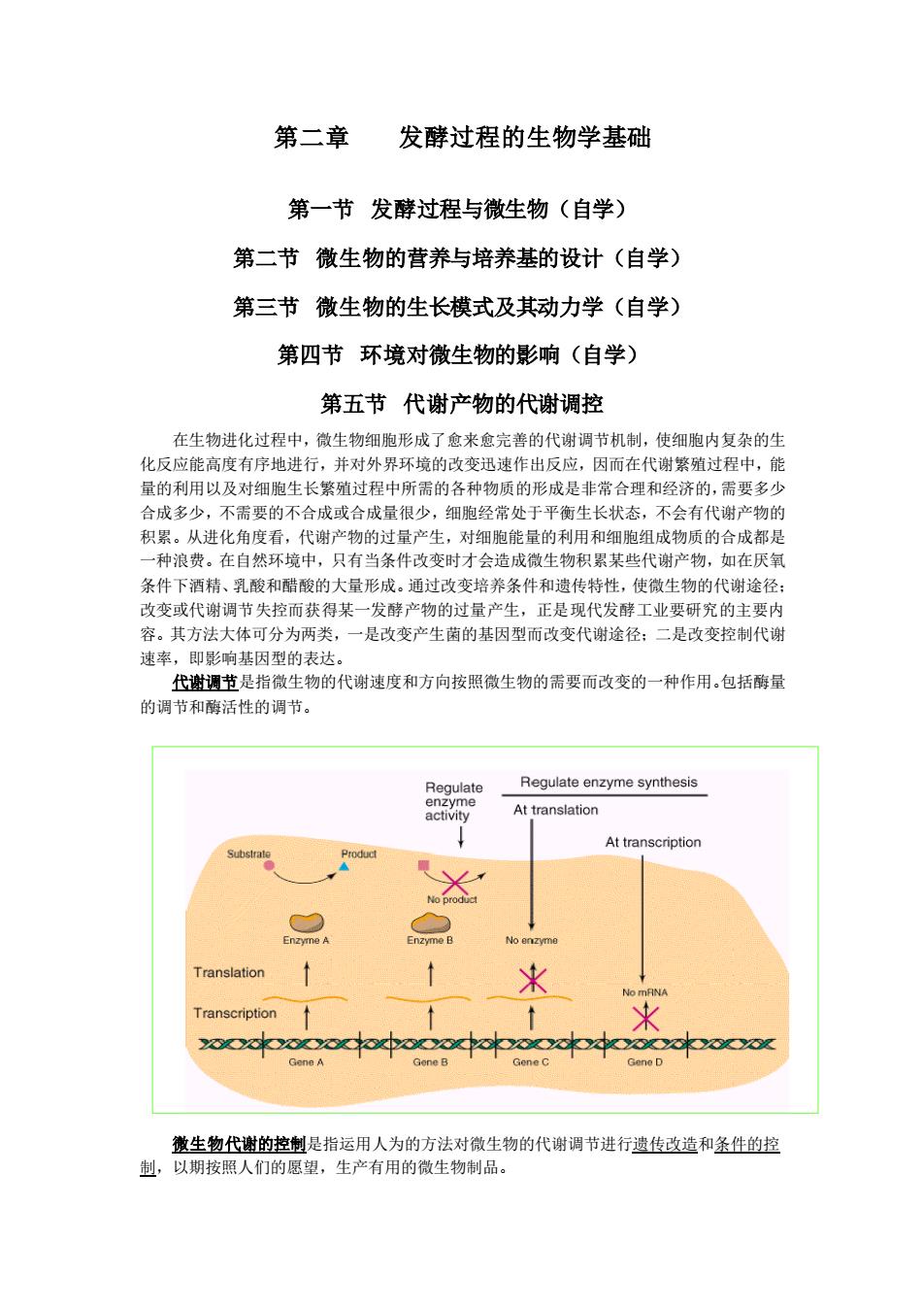
第二章 发酵过程的生物学基础 第一节 发酵过程与微生物(自学) 第二节 微生物的营养与培养基的设计(自学) 第三节 微生物的生长模式及其动力学(自学) 第四节 环境对微生物的影响(自学) 第五节 代谢产物的代谢调控 在生物进化过程中,微生物细胞形成了愈来愈完善的代谢调节机制,使细胞内复杂的生 化反应能高度有序地进行,并对外界环境的改变迅速作出反应,因而在代谢繁殖过程中,能 量的利用以及对细胞生长繁殖过程中所需的各种物质的形成是非常合理和经济的,需要多少 合成多少,不需要的不合成或合成量很少,细胞经常处于平衡生长状态,不会有代谢产物的 积累。从进化角度看,代谢产物的过量产生,对细胞能量的利用和细胞组成物质的合成都是 一种浪费。在自然环境中,只有当条件改变时才会造成微生物积累某些代谢产物,如在厌氧 条件下酒精、乳酸和醋酸的大量形成。通过改变培养条件和遗传特性,使微生物的代谢途径; 改变或代谢调节失控而获得某一发酵产物的过量产生,正是现代发酵工业要研究的主要内 容。其方法大体可分为两类,一是改变产生菌的基因型而改变代谢途径;二是改变控制代谢 速率,即影响基因型的表达。 代谢调节是指微生物的代谢速度和方向按照微生物的需要而改变的一种作用。包括酶量 的调节和酶活性的调节。 微生物代谢的控制是指运用人为的方法对微生物的代谢调节进行遗传改造和条件的控 制,以期按照人们的愿望,生产有用的微生物制品
第二章 发酵过程的生物学基础 第一节 发酵过程与微生物(自学) 第二节 微生物的营养与培养基的设计(自学) 第三节 微生物的生长模式及其动力学(自学) 第四节 环境对微生物的影响(自学) 第五节 代谢产物的代谢调控 在生物进化过程中,微生物细胞形成了愈来愈完善的代谢调节机制,使细胞内复杂的生 化反应能高度有序地进行,并对外界环境的改变迅速作出反应,因而在代谢繁殖过程中,能 量的利用以及对细胞生长繁殖过程中所需的各种物质的形成是非常合理和经济的,需要多少 合成多少,不需要的不合成或合成量很少,细胞经常处于平衡生长状态,不会有代谢产物的 积累。从进化角度看,代谢产物的过量产生,对细胞能量的利用和细胞组成物质的合成都是 一种浪费。在自然环境中,只有当条件改变时才会造成微生物积累某些代谢产物,如在厌氧 条件下酒精、乳酸和醋酸的大量形成。通过改变培养条件和遗传特性,使微生物的代谢途径; 改变或代谢调节失控而获得某一发酵产物的过量产生,正是现代发酵工业要研究的主要内 容。其方法大体可分为两类,一是改变产生菌的基因型而改变代谢途径;二是改变控制代谢 速率,即影响基因型的表达。 代谢调节是指微生物的代谢速度和方向按照微生物的需要而改变的一种作用。包括酶量 的调节和酶活性的调节。 微生物代谢的控制是指运用人为的方法对微生物的代谢调节进行遗传改造和条件的控 制,以期按照人们的愿望,生产有用的微生物制品

一、代谢调节方式 1,细胞透性的调节 细胞质膜的透性直接影响物质的吸收和代谢产物的分泌,从而影响到细胞内代谢的变 化。细胞质膜的透性的调节是微生物代谢调节的重要方式,由它控制着营养物质的吸收。 例如,大肠杆菌和鼠伤寒沙门氏菌吸收乳糖是由渗透酶和环状 AMP(cAMP)协同控制来 完成的。cAMP 的浓度是由腺苷酸环化酶(AC)的活性控制的,也就是说,乳糖的吸收受渗透 酶和 AMP 环化酶的控制,调节蛋白通过磷酸化的形式和腺苷酸环化酶(AC)或渗透酶结 合,分别使腺苷酸环化酶活化或使渗透酶失活。当有葡萄糖时,乳糖的渗透酶以无活性状态 存在,而腺苷酸环化酶也以非活性状态存在。 2,代谢途径区域化 原核微生物细胞结构虽然简单,但也划分出不同的区域,对于某一代谢途径有关的酶系 则集中某一区域,以保证这一代谢途径的酶促反应顺利进行,避免了其他途径的干扰。例如 呼吸的酶系集中在细胞质膜上;而与蛋白质合成有关的酶系则位于核蛋白体上;分解大分子 的水解酶,在革兰氏阴性菌里是位于壁膜间隙中,而革兰氏阳性菌则将这些水解酶类,分泌 于胞外。 在真核微生物细胞里,各种酶系被细胞器隔离分布。如与呼吸产能有关的酶系集中于线 粒体内膜上;蛋白质的合成酶系位于核蛋白体上;DNA 合成的某些酶位于细胞核里。 细胞具有复杂的膜结构使其代谢活动只能在特定的部位上进行,即代谢活动是区域化 的,其实质是控制酶与底物接触,使各个反应有序地进行。3,代谢流向的调控微生物在不 同条件下可以通过控制各代谢途径中某个酶促反应的速率来控制代谢物的流向,从而保持机 体代谢的平衡。它包括两种形式:由一个关键酶控制的可逆反应和由两种酶控制的逆单向反 应。(1)由一个关键酶控制的可逆反应 同一个酶可以通过不同辅基(或辅酶)控制代谢物的流向。例如,谷氨酸脱氢酶以 NADP+ 为辅酶时,主要是催化谷氨酸的合成,当以 NAD+为辅酶时,则催化谷氨酸的分解。因此微 生物可以通过不同的辅基来控制代谢物的流向。(2)由两种酶控制的逆单向反应 逆单向反应是在生物体代谢的关键部位的某些反应,它是由两种各自不同的酶来催化 的。即在一个“可逆”反应中,其中一种酶催化正反应,而另一种酶则催化逆反应。例如,葡 萄糖转化为 6-磷酸葡萄糖是由己糖激酶催化的,而其逆反应则是由 6-磷酸葡萄糖酯酶催化 的。6-磷酸果糖转化为 1,6-二磷酸果糖是由磷酸果糖激酶催化的,逆反应则由 1,6-二磷酸 果糖酯酶催化。4,代谢速度的调控在不可逆反应中,微生物通过调节酶的活性和酶量来控 制代谢物的流量。微生物在不同条件下能按照需要,通过酶活或抑制原有酶的活性或通过诱 导或阻遏酶的合成来自我调节其代谢速度,使之高度经济有效地利用能量和原科进行生长繁 殖。 二、酶合成的调节 1,酶合成的诱导 根据酶合成与底物的关系将酶分为组成型与诱导型两类。 组成酶是细胞固有的酶,其合成受相应基因控制,与底物、底物结构类似物及环境条件 无关,它主要用于调节初级代谢。 诱导酶是细胞为适应外来底物或底物结构类似物而临时合成的酶。如 E.coli 在含乳糖 培养基上产生的 β-半乳糖苷酶和半乳糖苷渗透酶就是由乳糖存在而诱导产生的。能促进诱 导酶产生的物质称为诱导物。底物、难以代谢的底物结构类似物及底物前体均可作为诱导物。 -酶合成的诱导分为协同诱导与顺序诱导两种类型。协同诱导指一种底物能同时诱导几种酶 的 合成。顺序诱导指先合成分解底物的酶,再依次合成分解各中间产物的酶,达到对复杂 代谢途径的分段调节。 2,酶合成的诱导的机制
一、代谢调节方式 1,细胞透性的调节 细胞质膜的透性直接影响物质的吸收和代谢产物的分泌,从而影响到细胞内代谢的变 化。细胞质膜的透性的调节是微生物代谢调节的重要方式,由它控制着营养物质的吸收。 例如,大肠杆菌和鼠伤寒沙门氏菌吸收乳糖是由渗透酶和环状 AMP(cAMP)协同控制来 完成的。cAMP 的浓度是由腺苷酸环化酶(AC)的活性控制的,也就是说,乳糖的吸收受渗透 酶和 AMP 环化酶的控制,调节蛋白通过磷酸化的形式和腺苷酸环化酶(AC)或渗透酶结 合,分别使腺苷酸环化酶活化或使渗透酶失活。当有葡萄糖时,乳糖的渗透酶以无活性状态 存在,而腺苷酸环化酶也以非活性状态存在。 2,代谢途径区域化 原核微生物细胞结构虽然简单,但也划分出不同的区域,对于某一代谢途径有关的酶系 则集中某一区域,以保证这一代谢途径的酶促反应顺利进行,避免了其他途径的干扰。例如 呼吸的酶系集中在细胞质膜上;而与蛋白质合成有关的酶系则位于核蛋白体上;分解大分子 的水解酶,在革兰氏阴性菌里是位于壁膜间隙中,而革兰氏阳性菌则将这些水解酶类,分泌 于胞外。 在真核微生物细胞里,各种酶系被细胞器隔离分布。如与呼吸产能有关的酶系集中于线 粒体内膜上;蛋白质的合成酶系位于核蛋白体上;DNA 合成的某些酶位于细胞核里。 细胞具有复杂的膜结构使其代谢活动只能在特定的部位上进行,即代谢活动是区域化 的,其实质是控制酶与底物接触,使各个反应有序地进行。3,代谢流向的调控微生物在不 同条件下可以通过控制各代谢途径中某个酶促反应的速率来控制代谢物的流向,从而保持机 体代谢的平衡。它包括两种形式:由一个关键酶控制的可逆反应和由两种酶控制的逆单向反 应。(1)由一个关键酶控制的可逆反应 同一个酶可以通过不同辅基(或辅酶)控制代谢物的流向。例如,谷氨酸脱氢酶以 NADP+ 为辅酶时,主要是催化谷氨酸的合成,当以 NAD+为辅酶时,则催化谷氨酸的分解。因此微 生物可以通过不同的辅基来控制代谢物的流向。(2)由两种酶控制的逆单向反应 逆单向反应是在生物体代谢的关键部位的某些反应,它是由两种各自不同的酶来催化 的。即在一个“可逆”反应中,其中一种酶催化正反应,而另一种酶则催化逆反应。例如,葡 萄糖转化为 6-磷酸葡萄糖是由己糖激酶催化的,而其逆反应则是由 6-磷酸葡萄糖酯酶催化 的。6-磷酸果糖转化为 1,6-二磷酸果糖是由磷酸果糖激酶催化的,逆反应则由 1,6-二磷酸 果糖酯酶催化。4,代谢速度的调控在不可逆反应中,微生物通过调节酶的活性和酶量来控 制代谢物的流量。微生物在不同条件下能按照需要,通过酶活或抑制原有酶的活性或通过诱 导或阻遏酶的合成来自我调节其代谢速度,使之高度经济有效地利用能量和原科进行生长繁 殖。 二、酶合成的调节 1,酶合成的诱导 根据酶合成与底物的关系将酶分为组成型与诱导型两类。 组成酶是细胞固有的酶,其合成受相应基因控制,与底物、底物结构类似物及环境条件 无关,它主要用于调节初级代谢。 诱导酶是细胞为适应外来底物或底物结构类似物而临时合成的酶。如 E.coli 在含乳糖 培养基上产生的 β-半乳糖苷酶和半乳糖苷渗透酶就是由乳糖存在而诱导产生的。能促进诱 导酶产生的物质称为诱导物。底物、难以代谢的底物结构类似物及底物前体均可作为诱导物。 -酶合成的诱导分为协同诱导与顺序诱导两种类型。协同诱导指一种底物能同时诱导几种酶 的 合成。顺序诱导指先合成分解底物的酶,再依次合成分解各中间产物的酶,达到对复杂 代谢途径的分段调节。 2,酶合成的诱导的机制

在没有诱导物存在时,调节基因 R 编码的阻遏蛋白与操纵基因 O 相结合,使附着于启动 基因 P 上的 RNA 聚合酶不能通过,从而阻止了 RNA 聚合酶对结构基因 S 的转录;当诱导物存 在时,阻遏蛋白因受诱导物作用而构型发生变化,失去与操纵基因的结合能力,从操纵基因 上解脱下来,使 RNA 聚合酶能对结构基因进行转录,进而翻译成酶蛋白。 3,酶合成的阻遏 在微生物的代谢过程中,当某途径的末端产物过量时,可通过阻碍该代谢途径中包括关 键酶在内的一系列酶的生物合成,彻底控制代谢和末端产物合成。阻遏作用有利于微生物从 合成源头节省有限的养料与能量。 (1)终产物阻遏 末端产物阻遏指某代谢途径末端产物过量累积引起的阻遏。在直线反应途径中,末端产 物阻遏较为简单,即产物作用于代谢途径中的各种酶,使这些酶不能合成。 (2)分解代谢产物阻遏 分解代谢物阻遏指细胞内同时存在两种底物(碳源或氮源)时,易利用底物会阻遏难利用 底物分解酶系的合成。其实质并非易利用底物直接导致,而是易利用底物分解过程中产生的 中间代谢物或末端代谢物的过量累积,阻遏了代谢途径中一些酶的合成。导致所谓“二次生 长现象”。 4,酶合成的阻遏的机制 终产物的反馈阻遏在转录水平上进行,终产物为辅阻遏物,它可激活由调节基因 R 生成 的无活性阻遏蛋白。辅阻遏物与阻遏蛋白结合形成活化阻遏物,它能与操纵基因 O 结合,阻 止 RNA 聚合酶对结构基因 S 的转录。 三、酶活性的调节
在没有诱导物存在时,调节基因 R 编码的阻遏蛋白与操纵基因 O 相结合,使附着于启动 基因 P 上的 RNA 聚合酶不能通过,从而阻止了 RNA 聚合酶对结构基因 S 的转录;当诱导物存 在时,阻遏蛋白因受诱导物作用而构型发生变化,失去与操纵基因的结合能力,从操纵基因 上解脱下来,使 RNA 聚合酶能对结构基因进行转录,进而翻译成酶蛋白。 3,酶合成的阻遏 在微生物的代谢过程中,当某途径的末端产物过量时,可通过阻碍该代谢途径中包括关 键酶在内的一系列酶的生物合成,彻底控制代谢和末端产物合成。阻遏作用有利于微生物从 合成源头节省有限的养料与能量。 (1)终产物阻遏 末端产物阻遏指某代谢途径末端产物过量累积引起的阻遏。在直线反应途径中,末端产 物阻遏较为简单,即产物作用于代谢途径中的各种酶,使这些酶不能合成。 (2)分解代谢产物阻遏 分解代谢物阻遏指细胞内同时存在两种底物(碳源或氮源)时,易利用底物会阻遏难利用 底物分解酶系的合成。其实质并非易利用底物直接导致,而是易利用底物分解过程中产生的 中间代谢物或末端代谢物的过量累积,阻遏了代谢途径中一些酶的合成。导致所谓“二次生 长现象”。 4,酶合成的阻遏的机制 终产物的反馈阻遏在转录水平上进行,终产物为辅阻遏物,它可激活由调节基因 R 生成 的无活性阻遏蛋白。辅阻遏物与阻遏蛋白结合形成活化阻遏物,它能与操纵基因 O 结合,阻 止 RNA 聚合酶对结构基因 S 的转录。 三、酶活性的调节

1,概念酶活性调节:是指一定数量的酶,通过其分子构象或分子结构的改变来调节其 催化反应的速率。 2,影响因素 影响酶活性的因素有: (1)底物和产物的性质和浓度 (2)环境因子(如压力、pH、离子强度和辅助因子等) (3)其他的酶的存在 3,调节方式 酶活性的调节方式有两种:激活已有酶的活性和抑制已有酶的活性 (1)激活激活:在激活剂的作用下,使原来无活性的酶变成有活性,或使原来活性低 的酶提高了活性的现象。 代谢调节的激活作用:主要是指代谢物对酶的激活。 前体激活,指代谢途径中后面的酶促反应,可被该途径中较前面的一个中间产物所促进。 代谢中间产物的反馈激活,指代谢中间产物对该代谢途径的前面的酶起激活作用(2) 抑制抑制和激活相反。由于某些物质的存在,降低酶活性,称为抑制。抑制可以是不可逆的, 这将造成代谢作用的停止;抑制也有可逆的,当抑制剂除去后,酶活性又恢复。在代谢调节 过程中所发生的抑制现象主要是可逆的,而且大多属于反馈抑制。 (2)反遗抑制:反馈抑制是指代谢的末端产物对酶(往往是代谢途径中的第一个酶)活 性的抑 制。反馈抑制作用在生物体内普通存在,它在维持细胞正常代谢、经济有效地利用 代谢原料、以及适应环境的变化,都具有重要作用。包括无分支代谢途径的调节和有分支代 谢途径的调节。 ①无分支代谢途径的调节通常是在线形的代谢途径中末端产物对催化第一步反应的酶 活性有抑制作用。例如,在大肠杆菌中,由苏氨酸(Thr)合成异亮氨酸(IIeu)时,异亮氨酸对 催化反应途径中的第一步反应的苏氨酸脱氨酶(TD)有抑制作用。 苏氨酸 α-酮丁酸 异亮氨酸
1,概念酶活性调节:是指一定数量的酶,通过其分子构象或分子结构的改变来调节其 催化反应的速率。 2,影响因素 影响酶活性的因素有: (1)底物和产物的性质和浓度 (2)环境因子(如压力、pH、离子强度和辅助因子等) (3)其他的酶的存在 3,调节方式 酶活性的调节方式有两种:激活已有酶的活性和抑制已有酶的活性 (1)激活激活:在激活剂的作用下,使原来无活性的酶变成有活性,或使原来活性低 的酶提高了活性的现象。 代谢调节的激活作用:主要是指代谢物对酶的激活。 前体激活,指代谢途径中后面的酶促反应,可被该途径中较前面的一个中间产物所促进。 代谢中间产物的反馈激活,指代谢中间产物对该代谢途径的前面的酶起激活作用(2) 抑制抑制和激活相反。由于某些物质的存在,降低酶活性,称为抑制。抑制可以是不可逆的, 这将造成代谢作用的停止;抑制也有可逆的,当抑制剂除去后,酶活性又恢复。在代谢调节 过程中所发生的抑制现象主要是可逆的,而且大多属于反馈抑制。 (2)反遗抑制:反馈抑制是指代谢的末端产物对酶(往往是代谢途径中的第一个酶)活 性的抑 制。反馈抑制作用在生物体内普通存在,它在维持细胞正常代谢、经济有效地利用 代谢原料、以及适应环境的变化,都具有重要作用。包括无分支代谢途径的调节和有分支代 谢途径的调节。 ①无分支代谢途径的调节通常是在线形的代谢途径中末端产物对催化第一步反应的酶 活性有抑制作用。例如,在大肠杆菌中,由苏氨酸(Thr)合成异亮氨酸(IIeu)时,异亮氨酸对 催化反应途径中的第一步反应的苏氨酸脱氨酶(TD)有抑制作用。 苏氨酸 α-酮丁酸 异亮氨酸
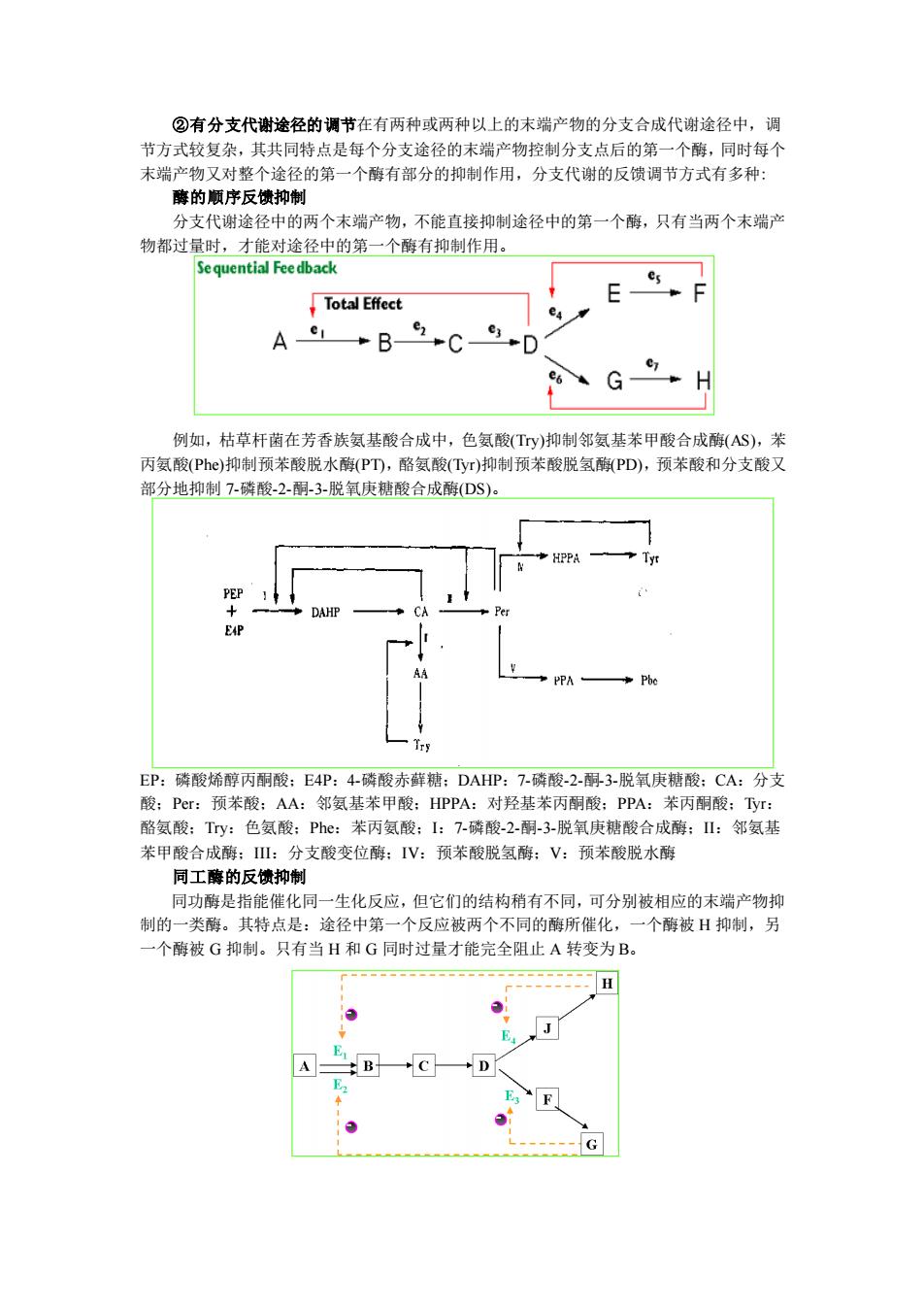
②有分支代谢途径的调节在有两种或两种以上的末端产物的分支合成代谢途径中,调 节方式较复杂,其共同特点是每个分支途径的末端产物控制分支点后的第一个酶,同时每个 末端产物又对整个途径的第一个酶有部分的抑制作用,分支代谢的反馈调节方式有多种: 酶的顺序反馈抑制 分支代谢途径中的两个末端产物,不能直接抑制途径中的第一个酶,只有当两个末端产 物都过量时,才能对途径中的第一个酶有抑制作用。 例如,枯草杆菌在芳香族氨基酸合成中,色氨酸(Try)抑制邻氨基苯甲酸合成酶(AS),苯 丙氨酸(Phe)抑制预苯酸脱水酶(PT),酪氨酸(Tyr)抑制预苯酸脱氢酶(PD),预苯酸和分支酸又 部分地抑制 7-磷酸-2-酮-3-脱氧庚糖酸合成酶(DS)。 EP:磷酸烯醇丙酮酸;E4P:4-磷酸赤藓糖;DAHP:7-磷酸-2-酮-3-脱氧庚糖酸;CA:分支 酸;Per:预苯酸;AA:邻氨基苯甲酸;HPPA:对羟基苯丙酮酸;PPA:苯丙酮酸;Tyr: 酪氨酸;Try:色氨酸;Phe:苯丙氨酸;I:7-磷酸-2-酮-3-脱氧庚糖酸合成酶;II:邻氨基 苯甲酸合成酶;III:分支酸变位酶;IV:预苯酸脱氢酶;V:预苯酸脱水酶 同工酶的反馈抑制 同功酶是指能催化同一生化反应,但它们的结构稍有不同,可分别被相应的末端产物抑 制的一类酶。其特点是:途径中第一个反应被两个不同的酶所催化,一个酶被 H 抑制,另 一个酶被 G 抑制。只有当 H 和 G 同时过量才能完全阻止 A 转变为 B
②有分支代谢途径的调节在有两种或两种以上的末端产物的分支合成代谢途径中,调 节方式较复杂,其共同特点是每个分支途径的末端产物控制分支点后的第一个酶,同时每个 末端产物又对整个途径的第一个酶有部分的抑制作用,分支代谢的反馈调节方式有多种: 酶的顺序反馈抑制 分支代谢途径中的两个末端产物,不能直接抑制途径中的第一个酶,只有当两个末端产 物都过量时,才能对途径中的第一个酶有抑制作用。 例如,枯草杆菌在芳香族氨基酸合成中,色氨酸(Try)抑制邻氨基苯甲酸合成酶(AS),苯 丙氨酸(Phe)抑制预苯酸脱水酶(PT),酪氨酸(Tyr)抑制预苯酸脱氢酶(PD),预苯酸和分支酸又 部分地抑制 7-磷酸-2-酮-3-脱氧庚糖酸合成酶(DS)。 EP:磷酸烯醇丙酮酸;E4P:4-磷酸赤藓糖;DAHP:7-磷酸-2-酮-3-脱氧庚糖酸;CA:分支 酸;Per:预苯酸;AA:邻氨基苯甲酸;HPPA:对羟基苯丙酮酸;PPA:苯丙酮酸;Tyr: 酪氨酸;Try:色氨酸;Phe:苯丙氨酸;I:7-磷酸-2-酮-3-脱氧庚糖酸合成酶;II:邻氨基 苯甲酸合成酶;III:分支酸变位酶;IV:预苯酸脱氢酶;V:预苯酸脱水酶 同工酶的反馈抑制 同功酶是指能催化同一生化反应,但它们的结构稍有不同,可分别被相应的末端产物抑 制的一类酶。其特点是:途径中第一个反应被两个不同的酶所催化,一个酶被 H 抑制,另 一个酶被 G 抑制。只有当 H 和 G 同时过量才能完全阻止 A 转变为 B