
第四章单克隆抗体 第四章单克隆抗体 单克隆抗体技术自1975年问世以来,在生命科学研究及临床医学诊断中发挥着越来越 重要的作用,每年与单克隆抗体相关的研究论文数以万计,生物产品数以千计,用于临床诊 断的单抗试剂日益增多。随着人类基因组计划的完成及蛋白质组计划的开始,单克隆抗体在 蛋白质功能研究、基因表达谱分析、临床疾病检测及治疗等方面的应用将更为广泛。本章主 要讨论鼠源性单克隆抗体制备的有关理论及在实际中经常碰到及需要注意的问题,并简述了 单克隆抗体技术的发展背景、原理和应用。单克隆抗体技术的问世,使免疫学研究迈向更高 和更精细的水平,单克隆抗体现已成为生物科学中最精细的探针之一。 第一节单克隆抗体的产生历史与原理 1975年Kohler和Milstein 以“分泌预定特异性抗体融合细胞的持续培养”(Continuous Culture of Fused Cells Secreting Antibody of Predefined Specificity)为题,在《自然》杂志首 次报道用仙台病毒使小鼠骨髓瘤细胞和经羊红细胞免疫的小鼠脾细胞融合产生的杂交瘤细 胞,既具有脾细胞分泌抗羊红细胞抗体的能力,又具有小鼠骨髓瘤细胞永生的特性,这一划 时代开创性工作翻开了人类利用杂交瘤抗体技术制备各类不同抗体广泛用于生命科学研究 及临床疾病诊断治疗的历史篇章,是生物技术发展史上的重要里程碑,Kohler和Milstein亦 因此获得1984年诺贝尔医学奖。 事实上在此之前,1970年Sinkovics等人已经报导过产生特异性病毒抗体的淋巴细胞和 由病毒引起的肿瘤细胞可以自然地在体内形成杂交瘤分泌特异性抗体。1973年Schwaber与 Coken首次报导了鼠一人杂交瘤的成功。1974年Bloom与Nakamura首次应用人的B细胞 与人的骨髓瘤细胞融合产生淋巴因子。以后于1980年Luben与Molle证明,在体外培养10 天的小鼠胸腺细胞能够产生淋巴因子,并能代替在体外培养中产生初次免疫(primary immunization,也称原发性免疫)反应所需要的免疫T细胞。他们应用这种胸腺细胞培养液 (称为条件培养液)加到小鼠脾细胞培养物中,并加入抗原(淋巴因子一破骨细胞激活因子) 刺激脾细胞产生免疫反应,随后用这种细胞与鼠骨髓瘤细胞杂交而产生破骨细胞激活因子的 单克隆抗体,建立了体外初次免疫反应,缩短了在体内免疫的时间。1978年Miller与Lipman 应用EB病毒转染人的B淋巴细胞产生单克隆抗体,使杂交瘤单克隆抗体技术向前迈进了一 步。 生物体产生抗体的最基本特点是抗体的多样性和不均质性,它们是针对众多不同抗原决 定簇(epitope)的混合性抗体。20世纪50年代以前对此现象一直没有满意的解释,1957年 Burnet创造性地提出了抗体产生的细胞系选择学说(即克隆选择学说),认为每个B细胞只 产生一种抗体,它以独特受体的形式存在于细胞表面,只能与一种抗原决定簇特异性地反应, 由此反应激活的这一淋巴细胞系,只能产生针对这一抗原决定簇的结构与功能完全相同的免 -49-
第四章 单克隆抗体 - 49 - 第四章 单克隆抗体 单克隆抗体技术自 1975 年问世以来,在生命科学研究及临床医学诊断中发挥着越来越 重要的作用,每年与单克隆抗体相关的研究论文数以万计,生物产品数以千计,用于临床诊 断的单抗试剂日益增多。随着人类基因组计划的完成及蛋白质组计划的开始,单克隆抗体在 蛋白质功能研究、基因表达谱分析、临床疾病检测及治疗等方面的应用将更为广泛。本章主 要讨论鼠源性单克隆抗体制备的有关理论及在实际中经常碰到及需要注意的问题,并简述了 单克隆抗体技术的发展背景、原理和应用。单克隆抗体技术的问世,使免疫学研究迈向更高 和更精细的水平,单克隆抗体现已成为生物科学中最精细的探针之一。 第一节 单克隆抗体的产生历史与原理 1975 年 Kohler 和 Milstein 以“分泌预定特异性抗体融合细胞的持续培养”(Continuous Culture of Fused Cells Secreting Antibody of Predefined Specificity)为题,在《自然》杂志首 次报道用仙台病毒使小鼠骨髓瘤细胞和经羊红细胞免疫的小鼠脾细胞融合产生的杂交瘤细 胞,既具有脾细胞分泌抗羊红细胞抗体的能力,又具有小鼠骨髓瘤细胞永生的特性,这一划 时代开创性工作翻开了人类利用杂交瘤抗体技术制备各类不同抗体广泛用于生命科学研究 及临床疾病诊断治疗的历史篇章,是生物技术发展史上的重要里程碑,Kohler 和 Milstein 亦 因此获得 1984 年诺贝尔医学奖。 事实上在此之前,1970 年 Sinkovics 等人已经报导过产生特异性病毒抗体的淋巴细胞和 由病毒引起的肿瘤细胞可以自然地在体内形成杂交瘤分泌特异性抗体。1973 年 Schwaber 与 Coken 首次报导了鼠-人杂交瘤的成功。1974 年 Bloom 与 Nakamura 首次应用人的 B 细胞 与人的骨髓瘤细胞融合产生淋巴因子。以后于 1980 年 Luben 与 Molle 证明,在体外培养 10 天的小鼠胸腺细胞能够产生淋巴因子,并能代替在体外培养中产生初次免疫(primary immunization,也称原发性免疫)反应所需要的免疫 T 细胞。他们应用这种胸腺细胞培养液 (称为条件培养液)加到小鼠脾细胞培养物中,并加入抗原(淋巴因子-破骨细胞激活因子) 刺激脾细胞产生免疫反应,随后用这种细胞与鼠骨髓瘤细胞杂交而产生破骨细胞激活因子的 单克隆抗体,建立了体外初次免疫反应,缩短了在体内免疫的时间。1978 年 Miller 与 Lipman 应用 EB 病毒转染人的 B 淋巴细胞产生单克隆抗体,使杂交瘤单克隆抗体技术向前迈进了一 步。 生物体产生抗体的最基本特点是抗体的多样性和不均质性,它们是针对众多不同抗原决 定簇(epitope)的混合性抗体。20 世纪 50 年代以前对此现象一直没有满意的解释,1957 年 Burnet 创造性地提出了抗体产生的细胞系选择学说(即克隆选择学说),认为每个 B 细胞只 产生一种抗体,它以独特受体的形式存在于细胞表面,只能与一种抗原决定簇特异性地反应, 由此反应激活的这一淋巴细胞系,只能产生针对这一抗原决定簇的结构与功能完全相同的免

第四章单克隆杭保 疫球蛋白(单克隆抗体)。随后,Nossal和edbers采用两种不同的抗原兔疫大白鼠,结果发 现,抗体生成细胞自始至终只能产生针对一个抗原的抗体,证明并支特了Bt的细胞系 选择学说。一个淋巴细胞只接受一个抗原决定族刺激及经刺激后扩增的淋巴细胞产生均一的 同质抗体,此即单克降抗体技术的理论基础。 制备单克隆抗体的基本过程是B细胞与骨髓瘤细胞的融合,在此基础上利用骨髓瘤细 胞的生化代谢缺陷在选择培养基中筛选培养,获得融合的杂交瘤细胞,因此,细胞融合和融 合细胞的筛选是杂交瘤抗体技术的基础,而理想的生化缺陷型骨髓瘤细胞系的建立是筛选融 合细胞的必需前提。一个浆细胞无限制增殖,导致产生一种均一的免疫球蛋白,称骨髓痴或 浆细胞瘤(plasmacytoma)。小鼠自发骨髓瘤极为罕见,需经人工诱导建立,1965年Sachs 等用矿物油刺激BALB/c小鼠诱生出骨髓瘤,并在体外培养成功,称为P3.1972年Milstein 等用8-杂氮鸟嘌呤(8-a2 guanine)对其进行诱导培养,从中筛选出缺乏次黄嘌吟.鸟嘌呤磷 酸核糖转移酶(hypoxanthine-guanine-,简称HGPRT)的新细胞系。 称为P3-X63-Ags。P3-X63-Ags因缺乏HGPRT而不能在HAT培养基中生长,但仍具分泌小 鼠1gGK轻链的能力,这显然不利于同B细胞融合后均一性抗体的产生。以后陆续诱导出的 一些新的突变株,如NS1,虽能合成K轻链,但无分泌能力,又如P3-X63-Ag8-653和SP20, 既不合成免疫球蛋白,也不分泌免疫球蛋白,从而保证了B细胞与骨髓瘤细胞融合后抗体 立生的均一性 细胞融合是杂交瘤抗体技术的基础。人们早就观察到在某些情况下或在体外培养中偶有 少数细胞可发生自发融合。早在1958年冈田等用高浓度的仙台病毒使小限艾氏腹水癌细胞 发生融合,其机制是病毒神经氨酸酶可降解细胞膜上的糖蛋白,使细胞膜局部凝集在病毒颗 粒的周用,在高pH及钙离子条件下,局部细胞质膜发生融合。1975年,Kao和Chavluk提 出用聚乙二醇作为细胞融合剂,其融合率较使用仙台病毒高出数百倍。目前己知的细胞融合 剂有数十种,而作为杂交瘤的融合剂主要是PG的行生物。EG是目前实验室制备杂交瘤 最常用的融合剂,其确切机制不清,可能因PEG的亲水性使细胞表面极性降低,导致脂双 层不稳定而引起细胞膜的融合。80年代末,Zimmermana报道了电融合技术,其原理是在电 场中沿电力线排列的细胞在高脉冲电场的作用下,细胞膜局部区域的双层脂分子结构遭到破 坏,出现微孔,细胞膜的通透性增加,继而产生相邻细胞的融合。电融合的融合率与PEG 相比有几何级数的增加,可减少B细胞的用量,如能利用一些亲和物质使B细胞与骨髓瘤 细胞在融合前配对接触,则可大大提高杂交细胞的产出率。 从上述不同技术及理论的发展历史可以看出,在20世纪70年代,免疫细胞的克隆选择 学说已被普遍接受,仙台病毒作为细胞融合剂已被成功使用,缺陷型小鼠骨髓瘤细胞系及融 合细胞的筛选方法均己建立,为单抗杂交瘤技术的出现奠定了理论及技术基础。1973年, Milstein等在研究抗体合成的遗传控制时,试图观察抗体合成的等位基因排斥规律能否在杂 交细胞上被打破。他们将大鼠的骨随瘤细胞和小鼠的骨随瘤细胞相融合,融合产生的杂交细 胞能分泌大鼠免疫球蛋白、小鼠免疫球蛋白和轻、重链分别来自大、小鼠两个亲代的杂交免 疫球蛋白,表明杂交细胞合成抗体不存在“等位基因排斥”现象,它们能共显地表达两个亲代 抗体信息。随后Kohler和Milstein用绵羊红细胞免疫小鼠,取脾淋巴细胞与小鼠骨随瘤细 胞P3-X63-Ag通过仙台病毒进行融合,然后于HAT培养基中选择培养,获得了能产生与绵 -50-
第四章 单克隆抗体 - 50 - 疫球蛋白(单克隆抗体)。随后,Nossal 和 edbers 采用两种不同的抗原免疫大白鼠,结果发 现,抗体生成细胞自始至终只能产生针对一个抗原的抗体,证明并支持了 Burnet 的细胞系 选择学说。一个淋巴细胞只接受一个抗原决定簇刺激及经刺激后扩增的淋巴细胞产生均一的 同质抗体,此即单克隆抗体技术的理论基础。 制备单克隆抗体的基本过程是 B 细胞与骨髓瘤细胞的融合,在此基础上利用骨髓瘤细 胞的生化代谢缺陷在选择培养基中筛选培养,获得融合的杂交瘤细胞,因此,细胞融合和融 合细胞的筛选是杂交瘤抗体技术的基础,而理想的生化缺陷型骨髓瘤细胞系的建立是筛选融 合细胞的必需前提。一个浆细胞无限制增殖,导致产生一种均一的免疫球蛋白,称骨髓瘤或 浆细胞瘤 (plasmacytoma)。小鼠自发骨髓瘤极为罕见,需经人工诱导建立,1965 年 Sachs 等用矿物油刺激 BALB/c 小鼠诱生出骨髓瘤,并在体外培养成功,称为 P3。1972 年 Milstein 等用 8-杂氮鸟嘌呤(8-azaguanine)对其进行诱导培养,从中筛选出缺乏次黄嘌呤-鸟嘌呤磷 酸核糖转移酶(hypoxanthine-guanine-phosphoribosyl transferase,简称 HGPRT)的新细胞系, 称为 P3-X63-Ags。P3-X63-Ags 因缺乏 HGPRT 而不能在 HAT 培养基中生长,但仍具分泌小 鼠 IgGκ轻链的能力,这显然不利于同 B 细胞融合后均一性抗体的产生。以后陆续诱导出的 一些新的突变株,如 NS1,虽能合成κ轻链,但无分泌能力,又如 P3-X63-Ag8-653 和 SP2/0, 既不合成免疫球蛋白,也不分泌免疫球蛋白,从而保证了 B 细胞与骨髓瘤细胞融合后抗体 产生的均一性。 细胞融合是杂交瘤抗体技术的基础。人们早就观察到在某些情况下或在体外培养中偶有 少数细胞可发生自发融合。早在 1958 年冈田等用高浓度的仙台病毒使小鼠艾氏腹水癌细胞 发生融合,其机制是病毒神经氨酸酶可降解细胞膜上的糖蛋白,使细胞膜局部凝集在病毒颗 粒的周围,在高 pH 及钙离子条件下,局部细胞质膜发生融合。1975 年,Kao 和 Chayluk 提 出用聚乙二醇作为细胞融合剂,其融合率较使用仙台病毒高出数百倍。目前已知的细胞融合 剂有数十种,而作为杂交瘤的融合剂主要是 PEG 的衍生物。PEG 是目前实验室制备杂交瘤 最常用的融合剂,其确切机制不清,可能因 PEG 的亲水性使细胞表面极性降低,导致脂双 层不稳定而引起细胞膜的融合。80 年代末,Zimmermana 报道了电融合技术,其原理是在电 场中沿电力线排列的细胞在高脉冲电场的作用下,细胞膜局部区域的双层脂分子结构遭到破 坏,出现微孔,细胞膜的通透性增加,继而产生相邻细胞的融合。电融合的融合率与 PEG 相比有几何级数的增加,可减少 B 细胞的用量,如能利用一些亲和物质使 B 细胞与骨髓瘤 细胞在融合前配对接触,则可大大提高杂交细胞的产出率。 从上述不同技术及理论的发展历史可以看出,在 20 世纪 70 年代,免疫细胞的克隆选择 学说已被普遍接受,仙台病毒作为细胞融合剂已被成功使用,缺陷型小鼠骨髓瘤细胞系及融 合细胞的筛选方法均已建立,为单抗杂交瘤技术的出现奠定了理论及技术基础。1973 年, Milstein 等在研究抗体合成的遗传控制时,试图观察抗体合成的等位基因排斥规律能否在杂 交细胞上被打破。他们将大鼠的骨髓瘤细胞和小鼠的骨髓瘤细胞相融合,融合产生的杂交细 胞能分泌大鼠免疫球蛋白、小鼠免疫球蛋白和轻、重链分别来自大、小鼠两个亲代的杂交免 疫球蛋白,表明杂交细胞合成抗体不存在“等位基因排斥”现象,它们能共显地表达两个亲代 抗体信息。随后 Kohler 和 Milstein 用绵羊红细胞免疫小鼠,取脾淋巴细胞与小鼠骨髓瘤细 胞 P3-X63-Ags 通过仙台病毒进行融合,然后于 HAT 培养基中选择培养,获得了能产生与绵

第四章单克隆杭体 羊红细胞特异结合的抗体的杂交细胞克隆。将这种杂交细胞注入同种小鼠腹腔内可形成肘 瘤,故称为杂交瘤。 单克隆抗体技术的主要优点:正如Milstein所说,首先,它是由一个独立的细胞克隆产 生的抗体,是一种十分明确的化学制品,而不是一种可随每个免疫动物,甚至同一动物不同 次免疫而变化的、不明确的非同质混合物,杂交细胞的永久培养可无限制地提供结构与性质 完全相同的单克降抗体:其次,该技术提供了用不纯抗原制备纯抗体的理想方法。因此单克 隆抗体技术一问世,就立即引起了生命科学研究领域的极大重视及广泛兴趣,相应技术迅速 在全球范围内得到了广泛的开展与应用,数以万计分泌单抗的杂交瘤细胞在实验室产生。 与此同时,杂交瘤技术也有了充分的发展,为适应临床应用的需要,杂交瘤细胞与B 淋巴细胞的隘合或杂交瘤细胞与杂交瘤细胞的融合以产生双功能抗体,鼠骨髓瘤细胞与人B 淋巴细胞的融合或人骨髓瘤细胞与人B淋巴细胞的融合以产生人单克降抗体的技术相继出 现。 1984年开始利用DNA重组技术改造鼠单克隆抗体使之人源化,以便降低在人体内应用 的免疫原性。 I989年出现的噬菌体展示抗体库(phage display antibody library)技术,使获得完全人 源的单克隆抗体成为可能。 而转基因小鼠(transgenetic mice)的出现把分子生物学技术与鼠单克隆抗体技术的结合 推向了顶峰,该技术将人抗体基因导入小鼠胚胎干细胞,使小鼠B淋巴细胞能产生人源抗 体,然后再经常规小鼠杂交瘤单克隆抗体技术制备人单克隆抗体,解决了用人B淋巴细胞 融合难以获得人单克隆抗体杂交瘤的难题。 毫无疑问,鼠单克隆抗体杂交瘤技术及在此基础上衍生出来的不同种类抗体制备技术, 在今后的生命科学研究中将发挥越来越多的作用。 图4-1是单克隆抗体的制备流程。 -51
第四章 单克隆抗体 - 51 - 羊红细胞特异结合的抗体的杂交细胞克隆。将这种杂交细胞注入同种小鼠腹腔内可形成肿 瘤,故称为杂交瘤。 单克隆抗体技术的主要优点:正如 Milstein 所说,首先,它是由一个独立的细胞克隆产 生的抗体,是一种十分明确的化学制品,而不是一种可随每个免疫动物,甚至同一动物不同 次免疫而变化的、不明确的非同质混合物,杂交细胞的永久培养可无限制地提供结构与性质 完全相同的单克隆抗体;其次,该技术提供了用不纯抗原制备纯抗体的理想方法。因此单克 隆抗体技术一问世,就立即引起了生命科学研究领域的极大重视及广泛兴趣,相应技术迅速 在全球范围内得到了广泛的开展与应用,数以万计分泌单抗的杂交瘤细胞在实验室产生。 与此同时,杂交瘤技术也有了充分的发展,为适应临床应用的需要,杂交瘤细胞与 B 淋巴细胞的融合或杂交瘤细胞与杂交瘤细胞的融合以产生双功能抗体,鼠骨髓瘤细胞与人 B 淋巴细胞的融合或人骨髓瘤细胞与人 B 淋巴细胞的融合以产生人单克隆抗体的技术相继出 现。 1984 年开始利用 DNA 重组技术改造鼠单克隆抗体使之人源化,以便降低在人体内应用 的免疫原性。 1989 年出现的噬菌体展示抗体库(phage display antibody library)技术,使获得完全人 源的单克隆抗体成为可能。 而转基因小鼠(transgenetic mice)的出现把分子生物学技术与鼠单克隆抗体技术的结合 推向了顶峰,该技术将人抗体基因导入小鼠胚胎干细胞,使小鼠 B 淋巴细胞能产生人源抗 体,然后再经常规小鼠杂交瘤单克隆抗体技术制备人单克隆抗体,解决了用人 B 淋巴细胞 融合难以获得人单克隆抗体杂交瘤的难题。 毫无疑问,鼠单克隆抗体杂交瘤技术及在此基础上衍生出来的不同种类抗体制备技术, 在今后的生命科学研究中将发挥越来越多的作用。 图 4-1 是单克隆抗体的制备流程
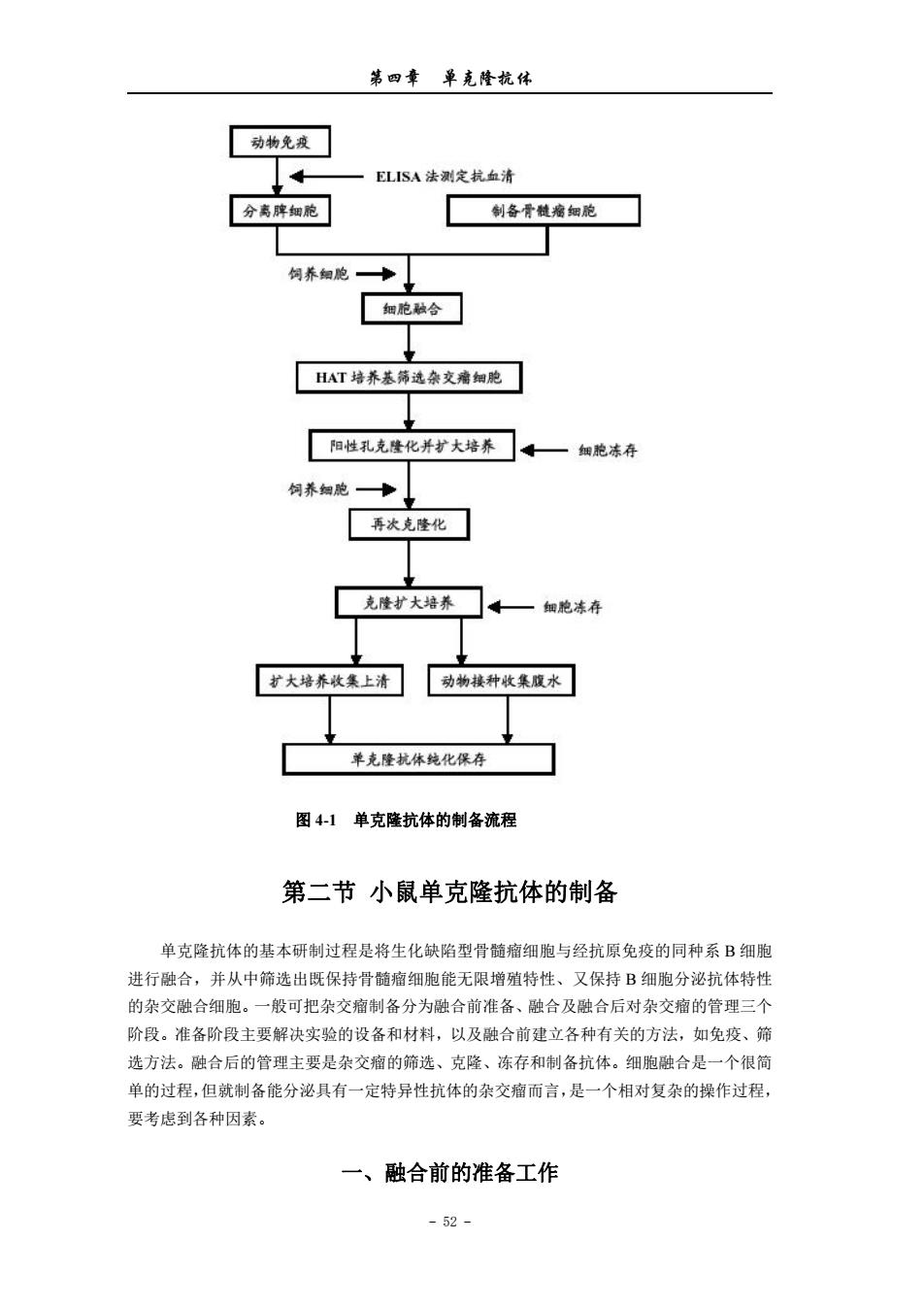
第四章单克隆杭体 动物免 ELISA法测定杭血清 分离脾细胞 制备骨髓瘤细胞 饲养细胞 HAT培养基选杂瘤细胞 阳性孔克隆化并扩大培养。一细胞冻行 饲养蜘胞—】 弄次克隆化 克隆扩大培养 扩大培养集上清动物楼种收集腹水 单克隆抗体纯化保存 图41单克隆抗体的制备流程 第二节小鼠单克隆抗体的制备 单克隆抗体的基本研制过程是将生化缺陷型骨髓痛细胞与经抗原免疫的同种系B细胞 进行融合,并从中筛选出既保持骨髓瘤细胞能无限增殖特性、又保持B细胞分泌抗体特性 的杂交融合细胞。一般可把杂交瘤制备分为融合前准备、融合及融合后对杂交瘤的管理三个 阶段。准备阶段主要解决实验的设备和材料,以及融合前建立各种有关的方法,如免疫、筛 选方法。融合后的管理主要是杂交瘤的筛选、克隆、冻存和制备抗体。细胞融合是一个很简 单的过程,但就制备能分泌具有一定特异性抗体的杂交瘤而言,是一个相对复杂的操作过程, 要考虑到各种因素。 一、融合前的准备工作 -52-
第四章 单克隆抗体 - 52 - 图 4-1 单克隆抗体的制备流程 第二节 小鼠单克隆抗体的制备 单克隆抗体的基本研制过程是将生化缺陷型骨髓瘤细胞与经抗原免疫的同种系 B 细胞 进行融合,并从中筛选出既保持骨髓瘤细胞能无限增殖特性、又保持 B 细胞分泌抗体特性 的杂交融合细胞。一般可把杂交瘤制备分为融合前准备、融合及融合后对杂交瘤的管理三个 阶段。准备阶段主要解决实验的设备和材料,以及融合前建立各种有关的方法,如免疫、筛 选方法。融合后的管理主要是杂交瘤的筛选、克隆、冻存和制备抗体。细胞融合是一个很简 单的过程,但就制备能分泌具有一定特异性抗体的杂交瘤而言,是一个相对复杂的操作过程, 要考虑到各种因素。 一、融合前的准备工作

第四章单克隆杭体 (一)组织培养材料的准备 1,主要设备 一般有组织培养能力的实验室均具有研制杂交瘤的基本条件。主要的仪器设备包括:超 净工作台、C02恒温培养箱、倒置光学显微镜、普通显微镜、离心机、液氨储存器、水浴恒 温装置、冰箱、培养液过滤装置、多孔培养板、塑料器皿及其它实验室常用的材料。 2.组织培养用水 一般都采用去离子水,将蒸馏水经超滤装置过滤后,即可除去其中的绝大部分离子。在 整个实验过程中都应该使用符合规格的用水。 3.培养基 制备单克隆抗体最常用的培养基有两种, 一种是改良的Dulbecco's Modified Eagle Medium(DMEM)。DMEM又有高糖(4500mgL)和低糖(1000mgL)两种。用于小鼠融 合的常为高糖DMEM。另一种为RPM-I64O。配制时干粉制剂搅全溶后,加水到需要的 体积,这样可以保证培养液的渗透压。经除菌过滤分装之后,即可保存于冰箱中,同时取少 量培养液进行培养证明其无茵,使用前根据需要再加入适量的牛血清、抗生素、碳酸氢钠、 丙酮酸盐和谷氨酰胺等成分,制备完全培养基所添加的各种成分都需要事先检菌。 4。血清 常用胎牛血清,但国内多数实验室常使用新生小牛血清,原因是新生小牛血清比胎牛血 清更便宜。不同来源及批号的血清对杂交瘤细胞的生长有明显差异。因此在实验前必须对不 同来源及批号的血清进行仔细的筛选检查,筛选出能支持单细胞生长或使细胞迅速增殖的血 清。在外观上稍有溶血而使血清中含有少量的血红蛋白并不影响其质量。常用的方法是用 株骨髓瘤细胞及两株杂交瘤细胞按有限稀释法(limited dilution)将细跑接种于若干个平行 孔,细胞培养液为含20%胎牛血清的DMEM或RPM-1640完全培养液,并于培养后3d、 7d、10d观察细胞增殖的情况,择优选用。 胎牛血清中球蛋白含量很低,一般不会影响杂交瘤细胞抗体分泌或抗体特异性的筛选。 此外胎生血清还含有一些生长刺激因子能促讲杂交瘤细胞的生长。如补充一些马血清,融合 细胞可迅速生长,同时又可节省一些胎牛血清。对于融合亲代细胞的培养一般采用含10% 胎牛血清培养基。但在融合、筛选及随后的克隆中一般用含20%的胎牛血清培养基。一旦 筛选出所需要的杂交瘤细胞后,可以将胎牛血清浓度逐步降低到10%或低至5%,使细胞缓 慢生长,以作为常规维持培养 根据需要在用前可以将胎牛血清灭活,即放在56℃水浴中温有30mi,以破坏血清中 补体的活性。也有报道在56℃加温45mn可以杀伤支原体。灭活前将在-20℃冻存的胎牛 血清取出,放在室温或4℃冰箱中待其自然解冻,将解冻的胎牛血清充分混匀后再放到56℃ 的恒温水浴中,温热30m,其间将血清瓶摇动混匀数次。将筛选灭活的牛血清或其它血清 分装成每次需要的数量,20℃保存。反复冻融会影响血清的质量。 5.抗生素 由于杂交瘤制备步骤多,持续培养的时间长,加之操作的误差和环境条件的不良常常会 -53
第四章 单克隆抗体 - 53 - (一) 组织培养材料的准备 1. 主要设备 一般有组织培养能力的实验室均具有研制杂交瘤的基本条件。主要的仪器设备包括:超 净工作台、CO2恒温培养箱、倒置光学显微镜、普通显微镜、离心机、液氮储存器、水浴恒 温装置、冰箱、培养液过滤装置、多孔培养板、塑料器皿及其它实验室常用的材料。 2. 组织培养用水 一般都采用去离子水,将蒸馏水经超滤装置过滤后,即可除去其中的绝大部分离子。在 整个实验过程中都应该使用符合规格的用水。 3. 培养基 制备单克隆抗体最常用的培养基有两种,一种是改良的 Dulbecco’s Modified Eagle Medium(DMEM)。DMEM 又有高糖(4500mg/L)和低糖(1000mg/L)两种。用于小鼠融 合的常为高糖 DMEM。另一种为 RPMI-1640。配制时干粉制剂搅拌全溶后,加水到需要的 体积,这样可以保证培养液的渗透压。经除菌过滤分装之后,即可保存于冰箱中,同时取少 量培养液进行培养证明其无菌,使用前根据需要再加入适量的牛血清、抗生素、碳酸氢钠、 丙酮酸盐和谷氨酰胺等成分,制备完全培养基所添加的各种成分都需要事先检菌。 4. 血清 常用胎牛血清,但国内多数实验室常使用新生小牛血清,原因是新生小牛血清比胎牛血 清更便宜。不同来源及批号的血清对杂交瘤细胞的生长有明显差异。因此在实验前必须对不 同来源及批号的血清进行仔细的筛选检查,筛选出能支持单细胞生长或使细胞迅速增殖的血 清。在外观上稍有溶血而使血清中含有少量的血红蛋白并不影响其质量。常用的方法是用一 株骨髓瘤细胞及两株杂交瘤细胞按有限稀释法(limited dilution)将细胞接种于若干个平行 孔,细胞培养液为含 20%胎牛血清的 DMEM 或 RPMI-1640 完全培养液,并于培养后 3d、 7d、10d 观察细胞增殖的情况,择优选用。 胎牛血清中球蛋白含量很低,一般不会影响杂交瘤细胞抗体分泌或抗体特异性的筛选。 此外胎牛血清还含有一些生长刺激因子能促进杂交瘤细胞的生长。如补充一些马血清,融合 细胞可迅速生长,同时又可节省一些胎牛血清。对于融合亲代细胞的培养一般采用含 10% 胎牛血清培养基。但在融合、筛选及随后的克隆中一般用含 20%的胎牛血清培养基。一旦 筛选出所需要的杂交瘤细胞后,可以将胎牛血清浓度逐步降低到 10%或低至 5%,使细胞缓 慢生长,以作为常规维持培养。 根据需要在用前可以将胎牛血清灭活,即放在 56℃水浴中温育 30 min,以破坏血清中 补体的活性。也有报道在 56 ℃加温 45 min 可以杀伤支原体。灭活前将在-20℃冻存的胎牛 血清取出,放在室温或 4℃冰箱中待其自然解冻,将解冻的胎牛血清充分混匀后再放到 56 ℃ 的恒温水浴中,温热 30 min,其间将血清瓶摇动混匀数次。将筛选灭活的牛血清或其它血清 分装成每次需要的数量,-20 ℃保存。反复冻融会影响血清的质量。 5. 抗生素 由于杂交瘤制备步骤多,持续培养的时间长,加之操作的误差和环境条件的不良常常会